Оптима кия клиренс: Kia Optima клиренс – Клиренс и дорожный просвет автомобилей
Меряем клиренс Киа Оптима
5.03.2017
Клиренс Киа Оптима — одна из характеристик, которыми часто интересуются покупатели этой машины. И неудивительно, дорожный просвет в немалой степени определяет возможности автомобиля. На проселочных, плохих, заснеженных дорогах, при проезде препятствий и объезде их по обочине, высокий автомобиль имеет преимущество в виде лучшей проходимости и защиты от повреждений. На ровных дорогах, наоборот нужен низкий клиренс, он делает центр тяжести ниже, что увеличивает устойчивость и управляемость машины. Высота дорожного просвета Киа зависит, в основном, от конструктивных особенностей, заложенных при проектировании, но ее можно изменить и самостоятельно используя тюнинговые детали подвески, резину с более высоким или низким профилем, проставки и другие элементы.
Реальные цифры клиренса Оптимы
Оптима относится к седанам D-класса и его клиренс вполне сопоставим с моделями этого класса. Данные о дорожном просвете Киа Оптима разнятся даже у дилеров. Некоторые заявляют цифру в 145 мм, у других в документации указывается 155 мм. В некоторых источниках можно встретить цифру в 160 мм. Так в чем же причина таких расхождений и какой клиренс у Киа Оптима на самом деле? Чтобы ответить на этот вопрос специально для читателей Iskorea.ru, автор этой статьи не поленился запастись рулеткой и отправиться к ближайшему дилеру Kia чтобы провести эксперимент. При замере клиренса Оптимы на 17-х колесах (других в наличии не оказалось) в самых низких местах дорожный просвет оказался равным примерно 155 мм. Такой же был указан и в разделе технических характеристик на сайте дилера. Откуда же берутся цифры в 145 и 160 см? Дело в том что высота клиренса зависит от высоты используемых шин. Кроме 17-дюймовых дисков на Киа Оптима, часто ставят 16 или 18 колеса. Это и дает небольшую разницу в дорожном просвете.
Некоторые заявляют цифру в 145 мм, у других в документации указывается 155 мм. В некоторых источниках можно встретить цифру в 160 мм. Так в чем же причина таких расхождений и какой клиренс у Киа Оптима на самом деле? Чтобы ответить на этот вопрос специально для читателей Iskorea.ru, автор этой статьи не поленился запастись рулеткой и отправиться к ближайшему дилеру Kia чтобы провести эксперимент. При замере клиренса Оптимы на 17-х колесах (других в наличии не оказалось) в самых низких местах дорожный просвет оказался равным примерно 155 мм. Такой же был указан и в разделе технических характеристик на сайте дилера. Откуда же берутся цифры в 145 и 160 см? Дело в том что высота клиренса зависит от высоты используемых шин. Кроме 17-дюймовых дисков на Киа Оптима, часто ставят 16 или 18 колеса. Это и дает небольшую разницу в дорожном просвете.
Клиренс в 155 мм можно назвать неплохим для седана в этом классе, хотя это вряд ли сыграет серьезную роль в его проходимости. Из-за достаточно длинной базы Оптима боится высоких неровностей и лежачих полицейских. Заезды на бордюр тоже не ее конек, да и крупные предметы тоже не стоит пропускать между колес. Но взамен всего этого автомобиль предлагает отличную управляемость, устойчивость на дороге и отсутствие кренов в поворотах.
Заезды на бордюр тоже не ее конек, да и крупные предметы тоже не стоит пропускать между колес. Но взамен всего этого автомобиль предлагает отличную управляемость, устойчивость на дороге и отсутствие кренов в поворотах.
Еще об автомобилях Optima:
Загрузка …Комментарии пользователей:
Kia Optima 2.4 GDI GT Line
| Название | 2.4 GDI GT Line |
| Класс | Бизнес-класс |
| Регион происхождения | Россия |
| Кузов | 4855х1860х1485х2805 |
| Двигатель | бензиновый 2.4, 188 л.с. |
| КПП | Автоматическая 6 |
| Привод | Передний |
| Расход топлива (город)(л/100км) | 12 |
| Расход топлива (шоссе)(л/100км) | 6.2 |
| Расход топлива (смешанный)(л/100км) | 8. 3 3 |
| Снаряженная масса (кг) | 1575 |
| Время разгона до 100 км/ч (сек.) | 9.1 |
| Макс.скорость (км/ч) | 210 |
| Количество подушек | 7 |
| Передняя ось | 235/45R18 |
| Задняя ось | 235/45R18 |
| Цена | 1 924 900 ₽ |
Безопасность |
| Передние подушки безопасности водителя и пассажира, боковые подушки безопасности, надувные занавески безопасности |
| ABS |
| ESP |
| Система помощи при трогании в гору |
| Система ЭраГлонасс |
| Система мониторинга давления в шинах |
| Светодиодные противотуманные фары |
| Светодиодные фары ближнего и дальнего света |
| Омыватели фар |
| Адаптивный свет |
| Подушка безопасности для коленей водителя |
| Система контроля слепых зон |
| Система помощи при выезде с парковки задним ходом |
| Система кругового обзора |
Автозвук и развлечения |
| Bluetooth-интерфейс |
| CD-магнитола с MP3 |
| AUX-разъем |
| USB-порт |
| Информационная система с цветным дисплеем |
| Навигационная система |
| Аудиосистема Hi-Fi класса |
Интерьер |
| Отделка руля и рычага КП кожей |
| Кожаный салон |
Внешний вид |
| Дневные ходовые огни светодиодные |
| Диски из легкого сплава |
| Панорамная стеклянная крыша |
| Люк с электроприводом |
Комфорт |
| Электропривод и обогрев зеркал |
| Электропривод складывания зеркал |
| Мультифункциональное рулевое колесо с лепестками переключения передач |
| Электростеклоподъемники передние и задние |
| Датчик света |
| Круиз-контроль |
| Обогрев лобового стекла в зоне покоя стеклоочистителей |
| Обогрев передних сидений |
| Многофункциональный цветной дисплей на приборной панели |
| Двухзонный климат-контроль |
| Обогрев руля |
| Датчик дождя |
| Электропривод сиденья водителя с памятью |
| Датчики парковки сзади и спереди |
| Доступ в салон и пуск двигателя без ключа |
| Электропривод двери багажника |
| Электрический стояночный тормоз |
| Камера заднего вида |
| Обогрев задних сидений |
| Беспроводное зарядное устройство |
| Электропривод сиденья пассажира |
| Вентиляция передних сидений |
| Система помощи при парковке |
Клиренс (дорожный просвет), мм Kia Optima
Дорожный просвет, он же клиренс очень важен покупателям, особенно покупателям в регионах.
Kia Optima продается в одной комплектации, поэтому клиренс всегда будет одинаковым, но не всегда. Иногда компании продают опции, увеличивающие клиренс и даже банальные колеса большего диаметра увеличат дорожный просвет Kia Optima. Дорожный просвет на Kia Optima всегда одинаков и равен 155 мм.
Для примера Клиренс (дорожный просвет), мм у конкурентов Kia Optima:
- Honda Accord – 130;
- Subaru Legacy – 150;
- Audi A4 – 140;
- Hyundai Sonata – 155;
- Mercedes-Benz C-Class – 130;
- Mazda 6 – 165;
Kia Optima
К обзору→| Комплектация | Клиренс (дорожный просвет), мм |
|---|---|
| 155 | |
2. 0 AT Специальная серия «Edition Plus» 0 AT Специальная серия «Edition Plus» | 155 |
| 2.0 AT Специальная серия «Лига Европы» | 155 |
| 2.0 AT Comfort | 155 |
| 2.0 AT Luxe | 155 |
| 2.0 AT Premium | 155 |
| 2.0 AT Prestige | 155 |
| 2.0 AT GT | 155 |
| 2.4 AT Специальная серия «Edition Plus» | 155 |
| 2.4 AT Специальная серия «Лига Европы» | 155 |
2. 4 AT GT Line 4 AT GT Line | 155 |
| 2.4 AT Luxe | 155 |
| 2.4 AT Prestige | 155 |
Клиренс (дорожный просвет), мм у конкурентов
Другие параметры Kia Optima
Размеры и вес автомобиля Kia Optima Мощность двигателя Kia Optima Максимальная скорость Kia Optima Крутящий момент двигателя Kia Optima Объем багажника автомобиля Kia Optima Объем двигателя Kia Optima Объем топливного бака Kia Optima Разгон до 100 км/ч Kia Optima Расход топлива Kia Optima Тип привода Kia Optima Коробка передач в Kia Optima Сколько мест в Kia Optima| Дорожный просвет | 145 мм |
| Количество мест | 5 |
| Высота | 1455 мм |
| Снаряженная масса | 1480 кг |
| Длина x Ширина x Высота | 4 845 x 1 830 x 1 455 мм |
| Колёсная база | 2795 мм |
| Допустимая полная масса | 2020 кг |
| Колея передних/задних колёс | 1 601/1 601 мм |
| Колея передних колёс | 1601 мм |
| Грузоподъёмность | 540 кг |
| Длина | 4845 мм |
| Объём багажника | 505 л |
| Колея задних колёс | 1601 мм |
| Минимальный объём багажника | 505 л |
| Ширина | 1830 мм |
| Обороты максимального крутящего момента | до 4 000 об/мин |
| Количество клапанов на цилиндр | 4 |
| Тип двигателя | Бензиновый |
| Объём двигателя | 2359 см3 |
| Наличие интеркулера | Нет |
| Диаметр цилиндра | |
| Конфигурация двигателя | Рядный |
| Мощность двигателя | 180 л.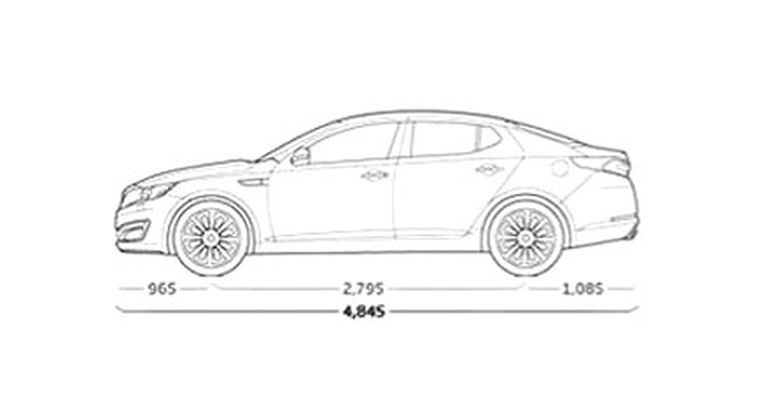 с. с. |
| Ход поршня | 97 мм |
| Тип впуска | Распределенный впрыск |
| Обороты максимальной мощности, макс. | 6000 об/мин |
| Максимальный крутящий момент | 231 Н•м |
| Обороты максимальной мощности | до 6 000 об/мин |
| Количество цилиндров | 4 |
| Обороты максимального крутящего момента, макс. | 4000 об/мин |
| Коробка передач | Автомат |
| Привод | Передний |
| Количество ступеней | 6 |
| Задние тормоза | Дисковые |
| Передняя подвеска | Независимая, пружинная, стойки макферсон, стабилизатор поперечной устойчивости |
| Задняя подвеска | Пружинная, независимая, амортизаторы, рычажная, стабилизатор поперечной устойчивости |
| Передние тормоза | Дисковые вентилируемые |
| Расход топлива в смешанном цикле | 8. 1 л/100 км 1 л/100 км |
| Время разгона до 100 км/ч | |
| Объём топливного бака | 70 л |
| Максимальная скорость | 210 км/ч |
| Запас хода | от 610 до 1 130 км |
| Рекомендуемое топливо | Аи-95 |
| Расход топлива в городе | 11.5 л/100 км |
| Экологический стандарт | Euro iv |
| Расход топлива на шоссе | 6.2 л/100 км |
| Диаметр разворота | 10.8 м |
| Усилитель руля | Электроусилитель |
| Ширина обода | 6,5 |
| Количество крепёжных отверстий | 5 |
| Диаметр расположения отверстий (PCD) | 114. 3 3 |
| Диаметр обода | 17 |
| Ширина обода | 6,5 |
| Количество крепёжных отверстий | 5 |
| Диаметр расположения отверстий (PCD) | 114.3 |
| Диаметр обода | 17 |
| Ширина профиля шины | 215 |
| Высота профиля шины | 55 |
| Диаметр шины | 17 |
| Высота профиля шины | 55 |
| Диаметр шины | 17 |
| Ширина профиля шины | 215 |
| Коробка передач | Автомат, 6 ст. |
| Привод | Передний |
| Диаметр разворота | 10. 8 м 8 м |
Кии и древки из углеродного волокна | Лучшие бильярдные кии
Стержни кия из углеродного волокна изменили бильярдный мир, как никакой другой продукт в новейшей истории. За годы исследований и разработок ведущие производители бильярдных киев открыли новые способы использования прочности легкого углеродного волокна для создания шафтов, которые не только прочны и долговечны, но и изгибаются там, где это необходимо для оптимального вращения и ощущения. Углеродное волокно аэрокосмического класса является сверхэффективным и обеспечивает наименьшую массу передней части, практически исключая разбрызгивание битка.Эти валы также заполнены специальной пеной, которая снижает вибрацию, обеспечивая баланс и постоянную обратную связь. Результатом является оборудование, спроектированное так, чтобы быть максимально производительным, наиболее стабильным и точным валом. Кроме того, они никогда не деформируются и не изнашиваются, поэтому вы можете рассматривать покупку как инвестиции в свою игру, которые окупятся на всю жизнь!
Кроме того, они никогда не деформируются и не изнашиваются, поэтому вы можете рассматривать покупку как инвестиции в свою игру, которые окупятся на всю жизнь!
Движение бильярдного кия из углеродного волокна не является маркетинговым заблуждением. Эти продукты представляют собой лучшее из лучшего и повсеместно используются ведущими игроками и профессионалами мира, включая Джона «Мр.626-дюймовый Шмидт, который смог превзойти Вилли Москони в хай-ране по пулу (удерживаемый более 65 лет) с его 12,4 Predator REVO .
Лучшие бренды кия из углеродного волокна:
Predator Cues находятся в авангарде технологии углеродного волокна и действительно положили начало движению, выпустив чрезвычайно популярную линейку Predator REVO. Валы Predator REVO доступны в размерах 12,4 мм и 12,9 мм и предлагаются с несколькими типами соединений.Они также выпустили брейк-стик из углеродного волокна, который используется в их брейк-киях BKRUSH. Они стали настолько популярными, что нам до сих пор трудно хранить их на складе спустя несколько лет после их первоначального запуска. Мы можем модернизировать любой из имеющихся у нас киев Predator, чтобы они поставлялись с древком REVO, доплатив разницу. Обычно это дополнительная плата в размере 190 долларов США. Если вам нужна помощь в совершении покупки с обновлением, позвоните нам.
Они стали настолько популярными, что нам до сих пор трудно хранить их на складе спустя несколько лет после их первоначального запуска. Мы можем модернизировать любой из имеющихся у нас киев Predator, чтобы они поставлялись с древком REVO, доплатив разницу. Обычно это дополнительная плата в размере 190 долларов США. Если вам нужна помощь в совершении покупки с обновлением, позвоните нам.
Cuetec выпустила свои реплики Cynergy Carbon Fibre с размером 12.Наконечник 5 мм с короткой белой визирной насадкой. Эти шафты, разработанные при участии чемпиона мира Шейна Ван Бонинга, известны своим хитом. Они доступны во всех основных стилях суставов и упакованы в классную специальную коробку с некоторыми дополнительными бесплатными вкусностями. Другие бренды, которые совершили скачок в углеродном волокне, — это Jacoby, Meucci, Miki Ignite (Mezz), у которых постоянно появляются новые варианты и стили суставов.
Техническое обслуживание вала из углеродного волокна:
Эти древки, естественно, очень гладкие и устойчивы к деформации, поэтому они не требуют особого ухода, но когда придет время, имейте в виду, что уход за ними отличается от того, как вы ухаживаете за деревянным древком. Во-первых, вы никогда не захотите использовать что-то абразивное. Обычно вы можете просто протереть их влажной тканью, чтобы очистить их. Некоторые производители рекомендуют использовать разбавленный медицинский спирт или мягкое средство для мытья посуды. Они также могут поставляться со специальными салфетками, которые вы можете использовать. Ознакомьтесь с рекомендациями производителя, прежде чем наносить что-либо на вал. Замена наконечника должна производиться авторизованным дилером или в ремонтном центре, чтобы предотвратить аннулирование гарантии. PoolDawg уполномочен всеми производителями предоставлять услуги по замене насадок.На большинство валов из углеродного волокна предоставляется пожизненная гарантия.
Во-первых, вы никогда не захотите использовать что-то абразивное. Обычно вы можете просто протереть их влажной тканью, чтобы очистить их. Некоторые производители рекомендуют использовать разбавленный медицинский спирт или мягкое средство для мытья посуды. Они также могут поставляться со специальными салфетками, которые вы можете использовать. Ознакомьтесь с рекомендациями производителя, прежде чем наносить что-либо на вал. Замена наконечника должна производиться авторизованным дилером или в ремонтном центре, чтобы предотвратить аннулирование гарантии. PoolDawg уполномочен всеми производителями предоставлять услуги по замене насадок.На большинство валов из углеродного волокна предоставляется пожизненная гарантия.
Если у вас есть какие-либо вопросы о покупке древков из углеродного волокна для вашего кия, подборе одного из них к новому прикладу, который мы носим, или какие-либо вопросы по обслуживанию, пожалуйста, обратитесь к нашим знающим сотрудникам, которые будут рады помочь !!! 866-843-3294
Киа | Корейский автопроизводитель присматривается к рынку кроссоверов B-класса с Stonic
Kia выпускает визуализацию дизайна совершенно нового Stonic, кроссовера B-сегмента, европейский дебют которого состоится в сентябре на автосалоне во Франкфурте.
На рендерах изображен кроссовер, мало чем отличающийся от варианта Rio Kia K2 Cross, который был показан в апреле на автосалоне в Шанхае и только что поступил в продажу в Китае.
«Stonic поступит в продажу на внутреннем корейском рынке и в Европе во второй половине 2017 года», — сообщил представитель WardsAuto . «Сроки запуска или планы для других рынков еще не подтверждены».
Представитель отказался сообщить, где был спроектирован и разработан Stonic или где будет производиться автомобиль, основанный на платформе Kia Rio.Платформа Rio четвертого поколения, выпущенного в прошлом году, была разработана Kia как в Германии, так и в Корее.
Поскольку Stonic использует ту же платформу, что и Rio, он, вероятно, находится в стадии серийного или предсерийного производства на заводе Kia в Сохари в Корее. Как и Rio, он, вероятно, будет производиться там и экспортироваться на европейский рынок.
Rio также производится на новом заводе Kia в Пескерии, Мексика, поэтому, если Kia Motors America решит продавать Stonic, он, скорее всего, будет включен в график производства Pesqueria вместе с Rio.
Внешний вид версии K2 Cross автомобиля Rio, который сейчас продается в Китае, во многом повторяет визуализацию, выпущенную для Stonic, особенно линия крыши, боковые панели кузова, большая часть передней части и даже поручни на крыше.
K2 Cross имеет ту же колесную базу, что и Rio, но листовой металл увеличивает его ширину на 1,2 дюйма. (30 мм) до 70 дюймов. (1750 мм). Длина в растянутом состоянии 1,6 дюйма. (40 мм) до 170 дюймов. (4240 мм). Дорожный просвет увеличен 1.8 дюймов (45 мм) больше, чем у Rio.
Stonic, как показано на рендеринге, имеет некоторую округлость, напоминающую Rio, но, как и K2 Cross, также, похоже, имеет дизайнерские черты новой Optima и роскошного спортивного гранд-турера Stinger.
Решетка радиатора Stonic в форме тигрового носа усовершенствована, чтобы придать новой модели спортивный и молодежный вид, который сохраняется под любым углом обзора спереди и сзади. Низкая покатая крыша имеет встроенный низкопрофильный багажник, очень похожий на багажник K2 Cross. Боковые панели кузова имеют гладкую форму ниже линии пояса. Дверные ручки, установленные заподлицо, выглядят так же, как и остальная часть автомобиля.
Боковые панели кузова имеют гладкую форму ниже линии пояса. Дверные ручки, установленные заподлицо, выглядят так же, как и остальная часть автомобиля.
Задняя часть Stonic имеет новую обработку боковых задних фонарей, которая, кажется, является первой для Kia. Как показано на изображениях, передний и задний бамперы, низко расположенная решетка радиатора, внутренние молдинги колесных арок и пороги имеют матово-черную отделку, которая эффектно контрастирует со смелым желтым верхним покрытием. Нижние молдинги с черной отделкой также характерны для K2 Cross, доступного в Китае.
Kia в своем официальном заявлении о выпуске намекает на множество высокотехнологичных опций и утверждает, что Stonic является «самой настраиваемой Kia из когда-либо созданных, внутри и снаружи». CUV будет иметь плавающий дисплей человеко-машинного интерфейса и панель управления.
В релизе также утверждается, что Stonic «привносит компактные размеры и низкий центр тяжести в B-класс», хотя, если показано, что он имеет те же размеры, что и K2 Cross, то китайский вариант Rio имеет избили его до отказа.
Некоторые наблюдатели предполагают, что Stonic будет предлагать те же двигатели, что и текущий Rio: 1,0-литровый 3-цил. турбо; турбодизель объемом 1,4 л; и 1,0 л, 1,25 л и 1,4 л, 4-цил. бензиновые версии. Представитель Kia отказывается комментировать возможную версию EV.
Несмотря на ориентированный на молодежь спортивный стиль, Stonic (и K2) выглядит более традиционно, чем его двоюродный брат, также только что представленный кроссовер Hyundai Kona B-класса. Kona будет официально представлен 13 июня в Сеуле вице-президентом Hyundai Чунг Ый-суном, сыном председателя Чунг Монг-Ку.
Возбуждающий интерес к Stonic не умаляет большого интереса во всем мире к тому, что будет раскрыто и сказано на следующей неделе о Kona. Но выпуск визуализаций — это напоминание Kia о том, что он тоже находится в игре CUV B-класса.
В своем пресс-релизе Kia называет название Stonic составным словом, составленным из части слова «скоростной» в сочетании с полным словом «тоник».
В музыке тоника — это первая и последняя нота музыкальной гаммы.Для более широкой публики тоник означает что-то, что поднимает настроение, что может быть лучшим объяснением.
Обзор: 2011 Kia Optima SX Turbo
Kia оправдывает свой слоган «способность удивлять» своей 274-сильной турбированной Optima SX.
На Юге какое-то движение, и ни о какой торнадной активности речи не идет.Kia приближается к финальной стадии полного редизайна своей продуктовой линейки, и новая Optima SX Turbo 2011 года занимает центральное место в возрождении бренда.
От залов заседаний в Сеуле, Южная Корея, до заводов на юге Соединенных Штатов, этот бренд находится на разрыве.
Используя опыт конструкторского отдела, возглавляемого Питером Шрейером, известным дизайнером Audi TT и Volkswagen New Beetle, фирма отбросила все остатки прошлого, как череду вредных привычек. Мы всегда в настроении попробовать новых вредных привычек. Присоединяйтесь, пока мы тестируем новую Optima Turbo.
Мы всегда в настроении попробовать новых вредных привычек. Присоединяйтесь, пока мы тестируем новую Optima Turbo.
Что это?
Пятиместный седан с тремя кузовами, эта Optima SX представляет собой усиленную версию автомобиля, который вызывал у нас восторг, когда мы ездили на нем в прошлом году.
Первый турбированный- Все, что когда-либо производила Kia (теперь к ним присоединился Sportage Turbo), он доступен в хорошо оснащенной модели EX с большим количеством гаджетов и плюшек.У нашей модели SX было еще больше.
Тесно связанный с Hyundai Sonata, Optima кажется немного более отточенным, как мы обсуждали несколько месяцев назад.
Что это против?
Сразу приходит на ум один конкурент: Hyundai Sonata Turbo. Помимо этого, внутри страны есть Ford Fusion, Toyota Camry и Nissan Altima, среди прочих.
Все достойные оппоненты, единственным определяющим фактором является то, любите ли вы пищу: мягкую, среднюю или острую. Хотя его корпоративный стабильный помощник, Sonata, имеет более плавный дизайн, Optima имеет тенденцию к заниженному, тихому виду.
Хотя его корпоративный стабильный помощник, Sonata, имеет более плавный дизайн, Optima имеет тенденцию к заниженному, тихому виду.
Есть прорывы?
Начните с версии 2.0T. Это двухточечные, как в турбо. Четырехцилиндровый двигатель с мощностью шести, это впечатляющий двигатель, который работает, когда вы наступаете этой кошке на хвост.
Продолжите с хорошо оснащенной моделью SX, в которой есть практически все, что входит в базовую комплектацию Optima. О, черт, давайте просто перейдем к делу и скажем, что этот автомобиль очень ценен.И все это можно получить по базовой цене, которая начинается ниже 26 долларов.
Как это выглядит?
Мы по-прежнему думаем, что Optima была красивой с самого начала. Длинный (190,1 дюйма), низкий (5,3 дюйма дорожного просвета) и широкий (72,1 дюйма) с пакетом SX он выглядит абсолютно по-тевтонски. Эта команда дизайнеров Шрейера, которая написала его, добавляет серьезности его уличному авторитету, моджо или чему-то еще. Но поставьте галочку напротив модели SX, и вы получите граунд-эффекты, высокопроизводительные сплавы в стиле Hot Wheel и многое другое.
Но поставьте галочку напротив модели SX, и вы получите граунд-эффекты, высокопроизводительные сплавы в стиле Hot Wheel и многое другое.
При внимательном рассмотрении автомобиля можно обнаружить множество элементов дизайна из (немецкого) Отечества, Азии и Скандинавии через Сеул. Начните с передних крыльев и боковых жабр, которые кажутся снятыми с BMW 5-Series последнего поколения. Добавьте дозу светодиодного освещения от Audi. Задний спойлер, вдохновленный Lexus IS, и задняя стойка, напоминающая нам о Saab 9-3. Вы не можете обвинить Kia в прямом воровстве, в отличие от вдохновения от вышеупомянутых автомобилей, но их выбор влияний, безусловно, впечатляет, а также очень вдохновляет.
А для любителей солнца опциональный панорамный (фактически двухпанельный) люк, который как раз оказался на нашем тестере.
А внутри?
Явно созданный для автолюбителей, Optima SX Turbo заимствует реплики у больших мальчиков, включая приборную панель, наклоненную к водителю. Подрулевые лепестки переключения передач входят в стандартную комплектацию нашего SX, как и сиденья с двухцветной обивкой из ткани и кожи. Отделка из искусственного углеродного волокна и алюминиевые педали поглощают усилия по внутренней отделке салона.
Подрулевые лепестки переключения передач входят в стандартную комплектацию нашего SX, как и сиденья с двухцветной обивкой из ткани и кожи. Отделка из искусственного углеродного волокна и алюминиевые педали поглощают усилия по внутренней отделке салона.
Оборудование высшего класса, включая окно водителя с автоматическим подъемом/опусканием, систему смарт-ключей и доступный технологический пакет. Это дает вам аудиосистему Infinity со спутниковым радио Sirius и Sirius Nav-Traffic, а также встроенную камеру обзора.
Kia использует девиз «Сила удивлять». порт подключения.
Материалы на шаг впереди Sonata, интересный шаг, учитывая, что Kia когда-то считалась младшим братом Hyundai. Какие сейчас отношения? Мы все еще не уверены.
Но идет ли?
Kia и дочерний бренд Hyundai находятся в новом ударе, чтобы покончить с прошлым, это сейчас. С этой целью они включают новый 2,0-литровый рядный четырехцилиндровый двигатель с непосредственным впрыском газа и турбинным компрессором для увеличения мощности с 200 лошадиных сил, как в базовом 2,4-литровом безнаддувном двигателе, до 274 л. .крутящего момента. Цифры EPA указаны как 22/34 — отлично, учитывая мощность под ногами. В среднем мы расходовали около 29 миль на галлон, что, учитывая вес нашей правой ноги на тонкой педали, является цифрой, с которой мы можем жить.
.крутящего момента. Цифры EPA указаны как 22/34 — отлично, учитывая мощность под ногами. В среднем мы расходовали около 29 миль на галлон, что, учитывая вес нашей правой ноги на тонкой педали, является цифрой, с которой мы можем жить.
О рядной четверке: Kia использовала турбокомпрессор с двойной спиралью, обеспечивающий наддув до 17,4 фунтов на квадратный дюйм. Лучшая часть этого уравнения — двухлопастная турбина, которая помогает минимизировать турбозадержку и подруливание крутящего момента. Kia добавила фронтальный интеркулер с новой системой Airguide, которая позволяет снизить температуру на выходе на 10 градусов.
Для простоты компания сократила выбор трансмиссии до одного: шестиступенчатая автоматическая коробка передач с функциями Sportmatic (manumatic), включая подрулевые лепестки переключения передач. Подобно тем, что были в Сонате, они позволяли нам играть в мальчика-гонщика на светофоре.
Подвеска со спортивной настройкой и южнокорейские автомобили обычно являются взаимоисключающими терминами. Но не здесь. Наша Optima оснащена парой стоек MacPherson спереди и многорычажным комплектом сзади.Стабилизаторы поперечной устойчивости и койловеры завершают детали и обеспечивают истинную езду и предлагают вдохновляющую управляемость для сегмента, который в последнее время не видел особого вдохновения. Наша неделя в Optima произвела на нас впечатление общей ездой, которая была менее плавной, чем у стандартной модели. Мы нашли место для руля, в отличие от чрезмерно усиленной версии, которую мы видели в двоюродной сестре Сонате.
Но не здесь. Наша Optima оснащена парой стоек MacPherson спереди и многорычажным комплектом сзади.Стабилизаторы поперечной устойчивости и койловеры завершают детали и обеспечивают истинную езду и предлагают вдохновляющую управляемость для сегмента, который в последнее время не видел особого вдохновения. Наша неделя в Optima произвела на нас впечатление общей ездой, которая была менее плавной, чем у стандартной модели. Мы нашли место для руля, в отличие от чрезмерно усиленной версии, которую мы видели в двоюродной сестре Сонате.
Дорожный шум на ровных дорогах обеспечил на удивление тихое вождение, даже с оптимизированным выхлопом нашей модели SX.Более грубые, композитные поверхности, как правило, немного повышали уровень шума. Мы просто хотим, чтобы Kia предложила шину, которая лучше справлялась с шумоподавлением, чем радиальные шины Nexen (корейской) марки, которыми была оснащена наша машина.
Недорогой угловой резак Optima удивлял нас практически на каждом шагу. Управляемость нашего тестового автомобиля обеспечила захватывающую езду, которая могла выявить Уолтера Митти в любом. Вентилируемые 12,6-дюймовые диски спереди и 11,2-дюймовые диски сзади быстро вернули нас к реальности.
Вентилируемые 12,6-дюймовые диски спереди и 11,2-дюймовые диски сзади быстро вернули нас к реальности.
Во время запуска Turbo Optim мы провели день на испытательном треке Leftlane , международной гоночной трассе Палм-Бич. В то время мы тестировали машину на трассе протяженностью 2,034 мили. Как бы ни было весело водить Optima, мы также пришли к выводу, что это не гоночная машина. Несомненно, в нем есть все забавные детали, из которых можно получить удовольствие от вождения, но в его текущей версии не садитесь в этот снегоход за 12 часов Себринга.
Но это должно подойти большинству из нас.
Почему вы его купите:
Потому что вам нравится заводить себе новые вредные привычки.
Почему бы и нет:
Потому что вы положили глаз на Camry ограниченной серии Frigidaire, которую вы видели в прошлые выходные в соседнем автоплексе.
Итоговый результат Leftlane
С мантрой «способность удивлять» Kia оставляет себя открытой для множества насмешек и презрения. Или нет. С Optima SX Turbo GDI у них есть соперник, который превосходит многих других в этом сегменте и предлагает множество вкусностей, которые вдохновляют всех нас на гонщиков.
Или нет. С Optima SX Turbo GDI у них есть соперник, который превосходит многих других в этом сегменте и предлагает множество вкусностей, которые вдохновляют всех нас на гонщиков.
Добавьте к этому 10-летнюю гарантию на 100 000 миль, и вы получите пакет, который удовлетворяет требованиям и менее наполняет.
Базовая цена Kia Optima SX Turbo 2011 года — 25 995 долларов. По результатам испытаний 30 840 долларов.
Технологический пакет, 2000 долларов США; Премиум-пакет SX, 2150 долларов США; Пункт назначения, 695 долларов.
Слова и фотографии Марка Элиаса.
границ | «Фагоцитарный синапс» и клиренс апоптотических клеток
Введение
Элиминация поврежденных или подвергнутых метаболическому стрессу клеток в многоклеточных организмах контролируется посредством включения апоптотических программ вместе с эффективными механизмами клиренса тканей (1-3).Взаимодействия фагоцитов/апоптотических клеток также инициируют противовоспалительное перепрограммирование, которое регулирует воспаление и иммунитет (4). Недостаточный клиренс апоптотических клеток способствует развитию и/или обострению многих аутоиммунных и воспалительных заболеваний [обзор в Ref. (5)].
Недостаточный клиренс апоптотических клеток способствует развитию и/или обострению многих аутоиммунных и воспалительных заболеваний [обзор в Ref. (5)].
Разнообразие молекулярных путей, опосредующих распознавание и фагоцитоз апоптотических клеток (эффероцитоз), отражает фундаментальную важность этого процесса (4). Существует несколько механизмов, с помощью которых жизнеспособные клетки избегают фагоцитоза (6).Однако измененный липидный состав плазматической мембраны (7, 8) и/или статус окисления (9) вместе с изменениями в репертуаре молекул клеточной поверхности и характером гликозилирования (10) называются «молекулярными паттернами, связанными с апоптотическими клетками» (11) (рис. 1). ), позволяют фагоцитам различать жизнеспособные и апоптотические клетки. Здесь мы рассматриваем формирование «эффероцитарного синапса» и сборку молекулярных платформ, которые облегчают фагоцитоз и последующие сигнальные события.
Рисунок 1 .Схематическое изображение поверхностных молекулярных изменений, связанных с апоптозом. Уменьшение экспрессии молекул клеточной поверхности может происходить в результате опосредованного металлопротеазами протеолитического отщепления. Сниженная экспрессия рецептора может ограничивать функцию апоптотических клеток и генерировать отличный от жизнеспособных клеток профиль клеточной поверхности. Например, сниженная экспрессия CD47 или отключенный CD31 могут привести к потере сигналов, препятствующих фагоцитозу. Кроме того, потеря регуляции интегрина может привести к функциональному разобщению в апоптотических клетках, что приводит к отслоению клеток.Измененная ассоциация молекул клеточной поверхности с липидными рафтами может изменить функциональную активность, включая усиление функции некоторых рецепторов, например, FcγRIIa на миелоидных клетках. Изменение переработки углеводов может привести к уменьшению воздействия сиаловой кислоты и появлению доступных остатков маннозы. Подавление регуляторных молекул комплемента (например, CD55 и CD46) может привести к опсонизации компонентами комплемента, включая C3b.
Уменьшение экспрессии молекул клеточной поверхности может происходить в результате опосредованного металлопротеазами протеолитического отщепления. Сниженная экспрессия рецептора может ограничивать функцию апоптотических клеток и генерировать отличный от жизнеспособных клеток профиль клеточной поверхности. Например, сниженная экспрессия CD47 или отключенный CD31 могут привести к потере сигналов, препятствующих фагоцитозу. Кроме того, потеря регуляции интегрина может привести к функциональному разобщению в апоптотических клетках, что приводит к отслоению клеток.Измененная ассоциация молекул клеточной поверхности с липидными рафтами может изменить функциональную активность, включая усиление функции некоторых рецепторов, например, FcγRIIa на миелоидных клетках. Изменение переработки углеводов может привести к уменьшению воздействия сиаловой кислоты и появлению доступных остатков маннозы. Подавление регуляторных молекул комплемента (например, CD55 и CD46) может привести к опсонизации компонентами комплемента, включая C3b. Воздействие анионных фосфолипидов, включая фосфатидилсерин (PtdSer), позволяет связывать широкий спектр опсонинов с апоптозными клетками.Белок S и Gas6 связываются с PtdSer зависимым от Ca 2+ образом, тогда как EGF-фактор 8 глобулы молочного жира (MFG-E8) связывается независимо от Ca 2+ . Другие менее четко определенные изменения апоптотической клеточной поверхности могут способствовать связыванию других опсонинов, включая тромбоспондин, С-реактивный белок и сурфактантный белок А. Наконец, на поверхности апоптотических клеток могут появляться белки с внутриклеточной локализацией, включая белки теплового шока и кальретикулин. Молекулы апоптотической клеточной поверхности заштрихованы синим, тогда как опсонины апоптотических клеток заштрихованы зеленым.
Воздействие анионных фосфолипидов, включая фосфатидилсерин (PtdSer), позволяет связывать широкий спектр опсонинов с апоптозными клетками.Белок S и Gas6 связываются с PtdSer зависимым от Ca 2+ образом, тогда как EGF-фактор 8 глобулы молочного жира (MFG-E8) связывается независимо от Ca 2+ . Другие менее четко определенные изменения апоптотической клеточной поверхности могут способствовать связыванию других опсонинов, включая тромбоспондин, С-реактивный белок и сурфактантный белок А. Наконец, на поверхности апоптотических клеток могут появляться белки с внутриклеточной локализацией, включая белки теплового шока и кальретикулин. Молекулы апоптотической клеточной поверхности заштрихованы синим, тогда как опсонины апоптотических клеток заштрихованы зеленым.
Фосфатидилсерин (PtdSer) в качестве лиганда для распознавания апоптотических клеток
Практически универсальным изменением мембраны, связанным с апоптозом, является каспаза-зависимая экспозиция PtdSer на внешнем листке плазматической мембраны (12–14) через XK-родственный белок 8 (15). Воздействие PtdSer влияет на биофизические характеристики и организацию плазматической мембраны за счет рекрутирования белков в обогащенные PtdSer области посредством электростатических взаимодействий (16).Фагоциты экспрессируют трансмембранные рецепторы, которые напрямую связывают PtdSer, например, специфичный для головного мозга ингибитор ангиогенеза-1 (BAI-1) (17) и стабилин-2 (18). Кроме того, растворимые молекулы, такие как транстиретиноподобный белок TTR-52 (19), глобула молочного жира EGF-фактор 8 (MFG-E8) (20, 21), белок S (Pros1), Gas6 (22) и C1q (23 ) также связываются (и опсонизируют) PtdSer, обеспечивая основу для распознавания фагоцитами через разнообразный набор контррецепторов.
Воздействие PtdSer влияет на биофизические характеристики и организацию плазматической мембраны за счет рекрутирования белков в обогащенные PtdSer области посредством электростатических взаимодействий (16).Фагоциты экспрессируют трансмембранные рецепторы, которые напрямую связывают PtdSer, например, специфичный для головного мозга ингибитор ангиогенеза-1 (BAI-1) (17) и стабилин-2 (18). Кроме того, растворимые молекулы, такие как транстиретиноподобный белок TTR-52 (19), глобула молочного жира EGF-фактор 8 (MFG-E8) (20, 21), белок S (Pros1), Gas6 (22) и C1q (23 ) также связываются (и опсонизируют) PtdSer, обеспечивая основу для распознавания фагоцитами через разнообразный набор контррецепторов.
Фагоциты не могут поглощать жизнеспособные клетки, которые экспонируют низкие уровни PtdSer во время активации (24–26) или когда экспозиция PtdSer индуцируется сверхэкспрессией фосфолипидной скрамблазы, трансмембранного белка 16F (TMEM16F) (27), что позволяет предположить, что необходимы дополнительные сигналы для инициировать эффероцитоз. Критический порог воздействия PtdSer на клеточную поверхность может быть необходим для запуска эффероцитоза (28). Например, распознавание PtdSer через Т-клеточный иммуноглобулин и молекулу, содержащую муциновый домен (TIM)-4, зависело от плотности лиганда, что позволяло фагоцитам различать высокий и низкий уровень воздействия PtdSer (28). Также могут иметь значение дальнейшие модификации PtdSer во время апоптоза, например, окисление или образование lyso-PtdSer (29).
Критический порог воздействия PtdSer на клеточную поверхность может быть необходим для запуска эффероцитоза (28). Например, распознавание PtdSer через Т-клеточный иммуноглобулин и молекулу, содержащую муциновый домен (TIM)-4, зависело от плотности лиганда, что позволяло фагоцитам различать высокий и низкий уровень воздействия PtdSer (28). Также могут иметь значение дальнейшие модификации PtdSer во время апоптоза, например, окисление или образование lyso-PtdSer (29).
Изменения рецепторов клеточной поверхности, связанные с апоптозом
Зависимая от апоптоза потеря рецепторов клеточной поверхности или появление «новых» молекул может способствовать распознаванию фагоцитами.Например, передача сигнала через сигнальный регуляторный белок -α (SIRPα) ингибирует миозин-II-опосредованный фагоцитоз (30). Предполагается, что подавление лигандов SIRPα, например, CD47 (31), с поверхности апоптотических клеток будет способствовать эффероцитозу (32, 33). Также сообщалось, что опосредованная SIRPα передача сигналов запускается связыванием сурфактантных белков (SP)-A и SP-D с фагоцитами. Однако SP-A может играть двойную роль в регуляции фагоцитоза, поскольку связывание с апоптотическими клетками/дебрисом приводит к стимулированию фагоцитоза через кальретикулин/CD91-опосредованный путь (34).Ранние эксперименты выявили уникальный чувствительный к заряду механизм распознавания апоптотических клеток (35). Клеточный гликокаликс обеспечивает «отталкивающую» силу отрицательного поверхностного заряда, которая противодействует межклеточным взаимодействиям (36). Потеря N-концевой сиаловой кислоты и воздействие фрагментов маннозы и фукозы во время апоптоза снижает электростатические силы, препятствующие распознаванию фагоцитов (37–39). Кроме того, поверхностный заряд апоптотических клеток дополнительно изменяется за счет снижения экспрессии сильно сиалилированных белков (например,г., CD43, CD45 и CD162) (37). В соответствии с этим предположением удаление сиаловой кислоты с клеточной поверхности при обработке нейраминидазой усиливает фагоцитоз (37, 39).
Однако SP-A может играть двойную роль в регуляции фагоцитоза, поскольку связывание с апоптотическими клетками/дебрисом приводит к стимулированию фагоцитоза через кальретикулин/CD91-опосредованный путь (34).Ранние эксперименты выявили уникальный чувствительный к заряду механизм распознавания апоптотических клеток (35). Клеточный гликокаликс обеспечивает «отталкивающую» силу отрицательного поверхностного заряда, которая противодействует межклеточным взаимодействиям (36). Потеря N-концевой сиаловой кислоты и воздействие фрагментов маннозы и фукозы во время апоптоза снижает электростатические силы, препятствующие распознаванию фагоцитов (37–39). Кроме того, поверхностный заряд апоптотических клеток дополнительно изменяется за счет снижения экспрессии сильно сиалилированных белков (например,г., CD43, CD45 и CD162) (37). В соответствии с этим предположением удаление сиаловой кислоты с клеточной поверхности при обработке нейраминидазой усиливает фагоцитоз (37, 39).
Апоптоз связан с потерей экспрессии регуляторных белков комплемента, таких как CD46/CD55 (40, 41). В результате может происходить отложение комплемента, что дает сигнал для распознавания фагоцитами. Дополнительные сигналы для фагоцитоза могут возникать в результате воздействия внутриклеточных белков, таких как кальретикулин (42) и аннексин I (43).После связывания с PtdSer на апоптотических клетках окисление Pros1 вызывает олигомеризацию, которая способствует Mer-зависимому фагоцитозу (44). Точно так же измененное гликозилирование мембранных белков или окисление липопротеиноподобных фрагментов низкой плотности на апоптотических клетках (8) также может способствовать специфическому распознаванию фагоцитами. Помимо повышенной экспрессии лигандов фагоцитарных рецепторов на апоптотических клетках, заплаты и/или кластеризация поверхностных молекул также могут иметь важные последствия для запуска фагоцитарного ответа.Кластеризация может происходить за счет специфической ассоциации с мембранными микродоменами. Например, FcγRIIa перераспределяется в микродомены мембраны во время апоптоза нейтрофилов (45).
В результате может происходить отложение комплемента, что дает сигнал для распознавания фагоцитами. Дополнительные сигналы для фагоцитоза могут возникать в результате воздействия внутриклеточных белков, таких как кальретикулин (42) и аннексин I (43).После связывания с PtdSer на апоптотических клетках окисление Pros1 вызывает олигомеризацию, которая способствует Mer-зависимому фагоцитозу (44). Точно так же измененное гликозилирование мембранных белков или окисление липопротеиноподобных фрагментов низкой плотности на апоптотических клетках (8) также может способствовать специфическому распознаванию фагоцитами. Помимо повышенной экспрессии лигандов фагоцитарных рецепторов на апоптотических клетках, заплаты и/или кластеризация поверхностных молекул также могут иметь важные последствия для запуска фагоцитарного ответа.Кластеризация может происходить за счет специфической ассоциации с мембранными микродоменами. Например, FcγRIIa перераспределяется в микродомены мембраны во время апоптоза нейтрофилов (45). Кроме того, специфический протеолиз молекул адгезии (например, CD62L) (46, 47) и несвязанная β 2 интегрин-опосредованная адгезия (47) во время апоптоза, вероятно, обеспечивают дополнительные молекулярные сигналы для фагоцитоза.
Кроме того, специфический протеолиз молекул адгезии (например, CD62L) (46, 47) и несвязанная β 2 интегрин-опосредованная адгезия (47) во время апоптоза, вероятно, обеспечивают дополнительные молекулярные сигналы для фагоцитоза.
Молекулы фагоцитов, опосредующие распознавание апоптотических клеток
Фагоциты способны к прямому распознаванию PtdSer, экспонированного на поверхности апоптотических клеток.BAI-1 связывается с PtdSer посредством повторов тромбоспондина (TSP) типа 1, присутствующих во внеклеточном домене (48). Связывание индуцирует образование тримерного комплекса BAI-1 с Rac-GEF ELMO и DOCK180, что способствует последующему поглощению апоптотических клеток (17, 49). Этот путь гомологичен генетически определенному пути удаления апоптотических клеток у Caenorhabditis elegans (Ced2-CrkII, Ced5-DOCK180, Ced10-Rac и Ced12-ELMO) (50).
Фосфатидилсерин также распознается семейством молекул CD300 с внеклеточным IgV-подобным доменом и сайтами связывания внутриклеточной адаптерной молекулы (51). CD300b локализуется в фагоцитарных чашечках и связывается с DAP12, активируя Syk и PI3K/Akt для стимуляции фагоцитоза (52). Стабилин-2 связывается с PtdSer и также не обладает прямой сигнальной активностью (18). Однако цитоплазматический домен стабилина-2 может взаимодействовать с GULP, облегчая фагоцитоз (53). GULP также связывается с мотивами NPxY, присутствующими в цитоплазматических доменах CD91/LRP (белок, связанный с рецептором липопротеинов низкой плотности) и рецептором-мусорщиком Ced-1 C. elegans (54). Напротив, TIM-4 обеспечивает Ca 2+ -зависимое PtdSer-зависимое распознавание апоптотических клеток, но не имеет внутриклеточного сигнального потенциала (55).Таким образом, TIM-4 функционирует совместно с другими рецепторами, запускающими интернализацию апоптотических клеток.
CD300b локализуется в фагоцитарных чашечках и связывается с DAP12, активируя Syk и PI3K/Akt для стимуляции фагоцитоза (52). Стабилин-2 связывается с PtdSer и также не обладает прямой сигнальной активностью (18). Однако цитоплазматический домен стабилина-2 может взаимодействовать с GULP, облегчая фагоцитоз (53). GULP также связывается с мотивами NPxY, присутствующими в цитоплазматических доменах CD91/LRP (белок, связанный с рецептором липопротеинов низкой плотности) и рецептором-мусорщиком Ced-1 C. elegans (54). Напротив, TIM-4 обеспечивает Ca 2+ -зависимое PtdSer-зависимое распознавание апоптотических клеток, но не имеет внутриклеточного сигнального потенциала (55).Таким образом, TIM-4 функционирует совместно с другими рецепторами, запускающими интернализацию апоптотических клеток.
Непрямое распознавание апоптотических клеток фагоцитами также достигается за счет рецепторов фагоцитов, которые связываются с растворимыми опсонинами апоптотических клеток. В C. elegans , TTR-52 соединяет PtdSer, подвергшийся воздействию апоптотических клеток, с фагоцитом Ced-1 (19), который вместе с Ced-6 инициирует быстрое и эффективное поглощение трупов апоптотических клеток соседними клетками. Этот модуль белков (Ced1-MEGF-10, Ced6-GULP и Ced7-ABCA1) был определен генетически в C.Элеганс (50). Pros1 и Gas6 содержат Gla-домен, который связывает PtdSer Ca 2+ -зависимым образом (22), соединяясь с тирозинкиназами рецептора Tyro3/Axl/Mer, которые сигнализируют об интернализации частиц посредством внутренней киназной активности (56). Напротив, MFG-E8 связывает PtdSer Са 2+ -независимым образом и образует мостики с интегринами фагоцитов α v β 3/5 посредством пептидных мотивов аргинин-глицин-аспарагиновой кислоты (RGD) в C1. и С2 домены (21).TSP-1 также соединяет апоптотические клетки через фагоцитарный интегрин α v β 3 и CD36 (57, 58).
elegans , TTR-52 соединяет PtdSer, подвергшийся воздействию апоптотических клеток, с фагоцитом Ced-1 (19), который вместе с Ced-6 инициирует быстрое и эффективное поглощение трупов апоптотических клеток соседними клетками. Этот модуль белков (Ced1-MEGF-10, Ced6-GULP и Ced7-ABCA1) был определен генетически в C.Элеганс (50). Pros1 и Gas6 содержат Gla-домен, который связывает PtdSer Ca 2+ -зависимым образом (22), соединяясь с тирозинкиназами рецептора Tyro3/Axl/Mer, которые сигнализируют об интернализации частиц посредством внутренней киназной активности (56). Напротив, MFG-E8 связывает PtdSer Са 2+ -независимым образом и образует мостики с интегринами фагоцитов α v β 3/5 посредством пептидных мотивов аргинин-глицин-аспарагиновой кислоты (RGD) в C1. и С2 домены (21).TSP-1 также соединяет апоптотические клетки через фагоцитарный интегрин α v β 3 и CD36 (57, 58).
Использование кооперативных рецепторов при фагоцитозе апоптотических клеток
Фагоциты в различных тканях или микроокружении экспрессируют различные репертуары эффероцитарных рецепторов. Неясно, использует ли одна фагоцитирующая клетка множественные пути рецепторов для распознавания и интернализации одной мишени апоптоза. Однако особые молекулярные потребности для захвата и последующей интернализации могут потребовать участия множества рецепторов (42).Кроме того, сложная топология апоптотических молекул клеточной поверхности и коопсонизация PtdSer с различными белками могут определять спектр вовлеченных путей передачи сигнала, контролируя интернализацию и последующие фагоцитарные ответы контекстно-зависимым образом.
Неясно, использует ли одна фагоцитирующая клетка множественные пути рецепторов для распознавания и интернализации одной мишени апоптоза. Однако особые молекулярные потребности для захвата и последующей интернализации могут потребовать участия множества рецепторов (42).Кроме того, сложная топология апоптотических молекул клеточной поверхности и коопсонизация PtdSer с различными белками могут определять спектр вовлеченных путей передачи сигнала, контролируя интернализацию и последующие фагоцитарные ответы контекстно-зависимым образом.
Прикрепление IgG-опсонизированных частиц к FcγR происходит при 4°C (59), тогда как интернализация требует реорганизации цитоскелета и метаболической активности (60). Точно так же апоптотические клетки также могут быть связаны фагоцитами через Mer при низкой температуре (61).Однако на авидность низкоаффинных рецепторов влияет плотность рецепторов и быстрое латеральное перемещение рецепторов для облегчения захвата мишени (62, 63). Подвижность рецепторов контролируется цитоскелетным ограничением, ассоциацией с липидными микродоменами мембраны и/или др.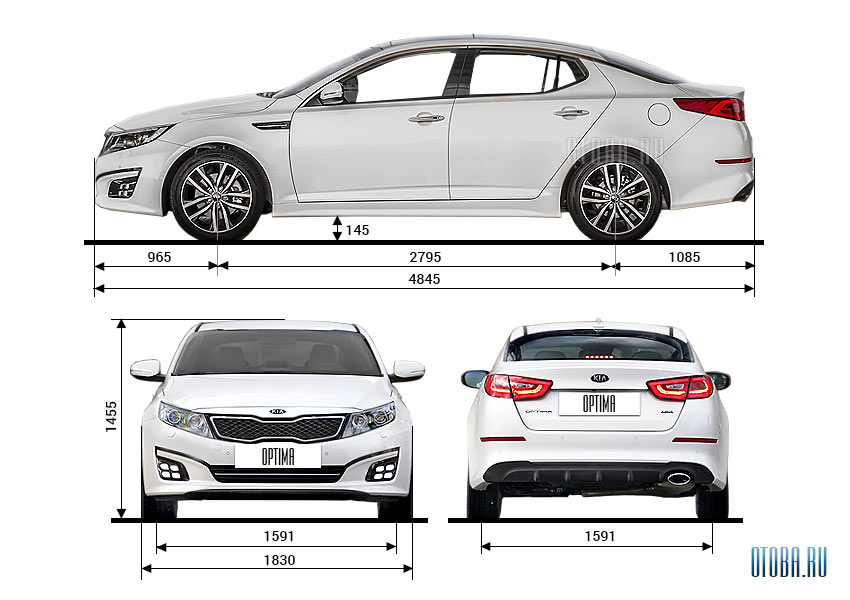 мембранными белками (64). Например, ассоциированный с цитоскелетом CD44 ограничивает подвижность мембранных липидов и рецепторов посредством взаимодействий с гиалуроновой кислотой, образуя гликозаминогликановый барьер, снижающий связывание фагоцитарных мишеней (36).Интересно, что перекрестное связывание CD44 с антителами усиливает макрофагальный фагоцитоз апоптотических клеток, возможно, в результате изменений в регуляции цитоскелета (65, 66). Фагоцитарные мишени связаны с динамическими расширениями фагоцитирующих клеток, включая филоподии и мембранные бороздки (67), а агрегация рецепторов необходима для координации изменений цитоскелета, необходимых для интернализации [см. (68) для всестороннего обзора регуляции цитоскелета в фагоцитарных синапсах].
мембранными белками (64). Например, ассоциированный с цитоскелетом CD44 ограничивает подвижность мембранных липидов и рецепторов посредством взаимодействий с гиалуроновой кислотой, образуя гликозаминогликановый барьер, снижающий связывание фагоцитарных мишеней (36).Интересно, что перекрестное связывание CD44 с антителами усиливает макрофагальный фагоцитоз апоптотических клеток, возможно, в результате изменений в регуляции цитоскелета (65, 66). Фагоцитарные мишени связаны с динамическими расширениями фагоцитирующих клеток, включая филоподии и мембранные бороздки (67), а агрегация рецепторов необходима для координации изменений цитоскелета, необходимых для интернализации [см. (68) для всестороннего обзора регуляции цитоскелета в фагоцитарных синапсах].
В отличие от IgG или комплемента, прикрепленных к компонентам микробной клеточной стенки, молекулы на поверхности апоптотических клеток могут проявлять неограниченную латеральную подвижность вследствие протеолитического расщепления актина во время апоптоза (69). Таким образом, могут существовать ключевые механистические различия между эффероцитозом и FcγR-зависимым фагоцитозом. Взаимодействие свободно подвижных молекул с фагоцитарными рецепторами может приводить к сборке микрокластеров рецепторов и значительно влиять на фагоцитоз (33). Альтернативно, опсонизация апоптотических клеток может привести к образованию неподвижных молекулярных комплексов [таких как аннексин V (70, 71)], которые способствуют перераспределению фагоцитарных рецепторов, необходимых для передачи сигналов интернализации.
Таким образом, могут существовать ключевые механистические различия между эффероцитозом и FcγR-зависимым фагоцитозом. Взаимодействие свободно подвижных молекул с фагоцитарными рецепторами может приводить к сборке микрокластеров рецепторов и значительно влиять на фагоцитоз (33). Альтернативно, опсонизация апоптотических клеток может привести к образованию неподвижных молекулярных комплексов [таких как аннексин V (70, 71)], которые способствуют перераспределению фагоцитарных рецепторов, необходимых для передачи сигналов интернализации.
Молекулярная сегрегация и образование фагоцитарного «синапса»
Клеточный контакт между фагоцитами и апоптозными клетками имеет параллели с контактами антигенспецифических Т- и В-клеток с антигенпрезентирующими клетками, что приводит к установлению иммунологического синапса (72).Эта специализированная зона межклеточного контакта стабилизирует адгезию и способствует эффективной молекулярной коммуникации после антиген-специфических взаимодействий. Ряд различных биофизических факторов (включая плотность рецепторов, сродство связывания лигандов, молекулярные размеры и взаимодействия с элементами цитоскелета) способствуют динамическому перераспределению адгезивных и сигнальных рецепторов в различных областях плазматической мембраны (73, 74).
Ряд различных биофизических факторов (включая плотность рецепторов, сродство связывания лигандов, молекулярные размеры и взаимодействия с элементами цитоскелета) способствуют динамическому перераспределению адгезивных и сигнальных рецепторов в различных областях плазматической мембраны (73, 74).
После контакта фагоцитов с поверхностями, покрытыми IgG, или с поддерживаемыми липидными бислоями образование нанокластеров FcγRII указывает на активационную организацию перераспределения рецепторов (75).Кластеры FcγR локализуются перед бороздками на расширяющихся псевдоногах с быстрым рекрутированием Syk на продвигающиеся псевдоподии и последующим ретроградным движением к центру клетки (76). PI3K совместно локализуется с актином вокруг кластеров FcγR, что свидетельствует о распространении сигнала от FcγR и согласуется с PI3K-зависимым контролем реаранжировок актинового цитоскелета (76).
Хотя контакт фагоцитов с апоптозными клетками не изучался с помощью методов визуализации с высоким разрешением, используемых во взаимодействиях Т-клеток и АПК, молекулярная сегрегация также может быть ключевой особенностью образования эффероцитарных синапсов (см. Таблицу 1).Например, исключение фосфатаз (например, CD45) из иммунного синапса является важным ранним событием в инициации опосредованного Т-клеточным рецептором фосфорилирования Zap70 и Lck (77). Изменения в распределении CD45 могут также представлять собой общую черту изменений мембран, которые контролируют сигнальные события, связанные с фагоцитозом. Рецептор лектина C-типа, содержащий β-глюкан, Dectin-1, опосредует распознавание и фагоцитоз дрожжевых частиц. Во время опосредованного Dectin-1 фагоцитоза исключение CD45 и протеинтирозинфосфатазы рецепторного типа CD148 из зарождающейся фагоцитарной чашечки (78) важно для передачи сигналов, связанных с интернализацией частиц (79).Перераспределение фосфатаз в мембране фагоцитов, вероятно, необходимо для интернализации апоптотических клеток.
Таблицу 1).Например, исключение фосфатаз (например, CD45) из иммунного синапса является важным ранним событием в инициации опосредованного Т-клеточным рецептором фосфорилирования Zap70 и Lck (77). Изменения в распределении CD45 могут также представлять собой общую черту изменений мембран, которые контролируют сигнальные события, связанные с фагоцитозом. Рецептор лектина C-типа, содержащий β-глюкан, Dectin-1, опосредует распознавание и фагоцитоз дрожжевых частиц. Во время опосредованного Dectin-1 фагоцитоза исключение CD45 и протеинтирозинфосфатазы рецепторного типа CD148 из зарождающейся фагоцитарной чашечки (78) важно для передачи сигналов, связанных с интернализацией частиц (79).Перераспределение фосфатаз в мембране фагоцитов, вероятно, необходимо для интернализации апоптотических клеток.
Таблица 1 . Сравнение формирования фагоцитарного синапса при FcγR-опосредованном и апоптотическом клеточном фагоцитозе.
Анализ молекулярной основы перераспределения CD45 после лигирования FcγR предполагает нелинейный характер с образованием барьера исключения, который ограничивает доступ CD45 к месту контакта (79). CD45 представляет собой относительно жесткую молекулу, которая простирается в осевом направлении от плазматической мембраны примерно на 20 нм (81).Подобно молекулярному перераспределению, которое происходит во время формирования иммунных синапсов, исключение из фагоцитарных синапсов зависело от осевых молекулярных размеров CD45. В экспериментах с использованием химерных конструкций, в которых внеклеточная часть CD45 была заменена либо CD43 (такой же длины), либо CD2 (более короткой длины), молекула CD43/CD45 исключалась из фагоцитарного синапса, тогда как CD2/CD45 — нет (79). Было показано, что потребность в интегринах и регуляции цитоскелета необходима для установления зоны исключения CD45, которая выходит за пределы слоя IgG (79).Эти результаты предполагают, что интегрин-опосредованный контакт между фагоцитами и фагоцитарными мишенями облегчает взаимодействие фагоцитарных рецепторов при низкой плотности лиганда или при связывании с более крупными частицами.
CD45 представляет собой относительно жесткую молекулу, которая простирается в осевом направлении от плазматической мембраны примерно на 20 нм (81).Подобно молекулярному перераспределению, которое происходит во время формирования иммунных синапсов, исключение из фагоцитарных синапсов зависело от осевых молекулярных размеров CD45. В экспериментах с использованием химерных конструкций, в которых внеклеточная часть CD45 была заменена либо CD43 (такой же длины), либо CD2 (более короткой длины), молекула CD43/CD45 исключалась из фагоцитарного синапса, тогда как CD2/CD45 — нет (79). Было показано, что потребность в интегринах и регуляции цитоскелета необходима для установления зоны исключения CD45, которая выходит за пределы слоя IgG (79).Эти результаты предполагают, что интегрин-опосредованный контакт между фагоцитами и фагоцитарными мишенями облегчает взаимодействие фагоцитарных рецепторов при низкой плотности лиганда или при связывании с более крупными частицами.
Интересно, что для распознавания апоптотических клеток было предложено, чтобы интегрины действовали совместно с другими рецепторами, опосредуя фагоцитоз. Например, интегрины α v и CD36 необходимы для распознавания TSP-1, связанного с апоптотическими клетками (58). Сходным образом, фагоцитоз апоптотических клеток через TIM-4 требует интегринов β 1 и активации интегрин-зависимой передачи сигналов с участием киназ семейства Src и FAK (82).Кроме того, было обнаружено, что взаимодействия между интегрином β 5 и стабилином-2 способствуют фагоцитозу апоптотических клеток (83). Сходным образом коэкспрессия α v β 5 с Mer увеличивала активацию Rac-1, регуляцию цитоскелета и фагоцитоз апоптотических клеток (84). Интегрины могут непосредственно опосредовать распознавание опсонинов апоптотических клеток, например, RGD-зависимое связывание MFG-E8 (21) или фибронектина (85). Однако фагоцитоз мишеней апоптоза усиливается после адгезии макрофагов к внеклеточному матриксу посредством интегринов β 1 (86) и нарушается после воздействия окисленных молекул внеклеточного матрикса (87).
Например, интегрины α v и CD36 необходимы для распознавания TSP-1, связанного с апоптотическими клетками (58). Сходным образом, фагоцитоз апоптотических клеток через TIM-4 требует интегринов β 1 и активации интегрин-зависимой передачи сигналов с участием киназ семейства Src и FAK (82).Кроме того, было обнаружено, что взаимодействия между интегрином β 5 и стабилином-2 способствуют фагоцитозу апоптотических клеток (83). Сходным образом коэкспрессия α v β 5 с Mer увеличивала активацию Rac-1, регуляцию цитоскелета и фагоцитоз апоптотических клеток (84). Интегрины могут непосредственно опосредовать распознавание опсонинов апоптотических клеток, например, RGD-зависимое связывание MFG-E8 (21) или фибронектина (85). Однако фагоцитоз мишеней апоптоза усиливается после адгезии макрофагов к внеклеточному матриксу посредством интегринов β 1 (86) и нарушается после воздействия окисленных молекул внеклеточного матрикса (87). Мы предполагаем, что, как и в случае FcγR-опосредованного фагоцитоза, передача сигналов интегрина может регулировать организацию цитоскелета и облегчать прикрепление/фагоцитоз апоптотических клеток с низким уровнем опсонизации (36).
Мы предполагаем, что, как и в случае FcγR-опосредованного фагоцитоза, передача сигналов интегрина может регулировать организацию цитоскелета и облегчать прикрепление/фагоцитоз апоптотических клеток с низким уровнем опсонизации (36).
Для макрофагоподобных клеток человека экспрессия CD47 ограничивает фагоцитоз опсонизированных IgG эритроцитов (30). CD47 связывается с фагоцитарным SIRPα, что приводит к рекрутированию в фагоцитарный синапс, снижению накопления немышечного миозина IIa и уровней фосфорилирования тирозина (30).Локализация SIRPα в месте контакта с клеткой будет привлекать ингибирующие тирозинфосфатазы, такие как SHP-1 , посредством мотивов ITIM, присутствующих в цитоплазматическом домене SIRPα (88). В отсутствие CD47 или когда CD47 был заблокирован антителом, фагоцитоз усиливался. Специфические мембранные рецепторы организованы в белковые островки в неактивированных Т-клетках, которые впоследствии сливаются в результате передачи сигналов, опосредованной Т-клеточными рецепторами (89). Мембранное распределение SIRPα и FcγR во время фагоцитоза было дополнительно проанализировано с использованием микроскопии сверхвысокого разрешения.Когда макрофаги высевали на предметные стекла, покрытые поли-1-лизином, наблюдались молекулярные кластеры, содержащие как FcγRI, так и SIRPα (75). Было обнаружено, что эти молекулы разделяются на дискретные нанокластеры, когда макрофаги высевают на IgG. Интересно, что IgG способствовал образованию концентрических колец FcγRI и FcγRII, при этом FcγRI перераспределялся быстрее (<10 мин). Сходные результаты были получены при взаимодействии макрофагов с IgG в липидном бислое на подложке. При включении рекомбинантного CD47 в липидный бислой на носителе сегрегация FcγR и SIRPα и образование концентрических колец FcγR блокировались.Таким образом, совместная локализация SIRPα и FcγR ингибирует клеточную активацию после лигирования FcγR, тогда как сегрегация этих двух молекул приводит к активации (75). Можно предположить, что эффероцитарные рецепторы также будут присутствовать в динамически регулируемых нанокластерах в мембране фагоцитов.
Мембранное распределение SIRPα и FcγR во время фагоцитоза было дополнительно проанализировано с использованием микроскопии сверхвысокого разрешения.Когда макрофаги высевали на предметные стекла, покрытые поли-1-лизином, наблюдались молекулярные кластеры, содержащие как FcγRI, так и SIRPα (75). Было обнаружено, что эти молекулы разделяются на дискретные нанокластеры, когда макрофаги высевают на IgG. Интересно, что IgG способствовал образованию концентрических колец FcγRI и FcγRII, при этом FcγRI перераспределялся быстрее (<10 мин). Сходные результаты были получены при взаимодействии макрофагов с IgG в липидном бислое на подложке. При включении рекомбинантного CD47 в липидный бислой на носителе сегрегация FcγR и SIRPα и образование концентрических колец FcγR блокировались.Таким образом, совместная локализация SIRPα и FcγR ингибирует клеточную активацию после лигирования FcγR, тогда как сегрегация этих двух молекул приводит к активации (75). Можно предположить, что эффероцитарные рецепторы также будут присутствовать в динамически регулируемых нанокластерах в мембране фагоцитов.
Молекулярные размеры и распознавание апоптотических клеток
Зависимое от размера перераспределение молекул внутри мембраны фагоцитов может представлять собой важный организующий принцип для сборки молекулярных платформ, которые необходимы для передачи сигналов об изменениях цитоскелета, необходимых для интернализации апоптотических клеток.Оценка молекулярных размеров рецепторов, участвующих в эффероцитозе, с использованием опубликованных структурных данных (51, 90-106) выявила значительные различия в осевых размерах (рис. 2А).
Рисунок 2 . Молекулярные размеры белков, участвующих в фагоцитозе апоптотических клеток. (A) Некоторые из основных семейств рецепторов, которые, как было установлено, играют роль в фагоцитозе апоптотических клеток, изображены приблизительно в масштабе, основанном на доступных опубликованных данных кристаллографии/ЯМР или криоэлектронной микроскопии для различных белковых доменов. .Одно из предположений состоит в том, что молекулы имеют относительно жесткую структуру, хотя возможно, что гибкость значительно изменит осевую длину. Имеются значительные различия в размерах опсонинов апоптотических клеток и в рецепторах, которые опосредуют связывание фосфатидилсерина (PtdSer) (прямо или косвенно). (B) Примеры пар рецептор-контррецептор показаны для иллюстрации вероятных различий в межмембранном «рабочем» расстоянии во время фагоцитоза, особенно для молекул, участвующих в распознавании PtdSer (показаны желтым цветом).Рецепторы фагоцитов показаны синим цветом, опсонины показаны зеленым цветом, а апоптотическая клеточная мембрана показана красным цветом.
Имеются значительные различия в размерах опсонинов апоптотических клеток и в рецепторах, которые опосредуют связывание фосфатидилсерина (PtdSer) (прямо или косвенно). (B) Примеры пар рецептор-контррецептор показаны для иллюстрации вероятных различий в межмембранном «рабочем» расстоянии во время фагоцитоза, особенно для молекул, участвующих в распознавании PtdSer (показаны желтым цветом).Рецепторы фагоцитов показаны синим цветом, опсонины показаны зеленым цветом, а апоптотическая клеточная мембрана показана красным цветом.
Во-первых, следует отметить, что CD300 (одиночный Ig-подобный домен) и TIM-4 (одиночный Ig-подобный домен с потенциально жесткой муциноподобной ножкой), которые опосредуют прямое распознавание PtdSer, по прогнозам охватывают относительно короткий межмембранный расстояние между фагоцитом и мишенью (4 и 7 нм соответственно, рис. 2В). Напротив, стабилин-2, который также связывается с PtdSer, представляет собой гораздо более длинную молекулу длиной около 23 нм. Предполагая некоторую структурную гибкость, BAI-1 может функционировать в широком диапазоне межмембранных расстояний. Внеклеточная область BAI-1 содержит 5 повторов TSP типа 1 (размером около 5 нм) с мотивами связывания LPS и PtdSer, которые вместе с областями GAIN / HBD могут простираться до ~ 33 нм от плазматической мембраны. После начального связывания PtdSer с N-концевым повтором TSP BAI-1 может располагаться параллельно апоптотической клеточной поверхности по мере лигирования дополнительных повторов TSP (рис. 2В).Тем не менее, представляется вероятным, что необходима определенная степень молекулярной сегрегации для того, чтобы эти разные рецепторы участвовали в апоптотическом захвате клеток одним и тем же фагоцитом.
Предполагая некоторую структурную гибкость, BAI-1 может функционировать в широком диапазоне межмембранных расстояний. Внеклеточная область BAI-1 содержит 5 повторов TSP типа 1 (размером около 5 нм) с мотивами связывания LPS и PtdSer, которые вместе с областями GAIN / HBD могут простираться до ~ 33 нм от плазматической мембраны. После начального связывания PtdSer с N-концевым повтором TSP BAI-1 может располагаться параллельно апоптотической клеточной поверхности по мере лигирования дополнительных повторов TSP (рис. 2В).Тем не менее, представляется вероятным, что необходима определенная степень молекулярной сегрегации для того, чтобы эти разные рецепторы участвовали в апоптотическом захвате клеток одним и тем же фагоцитом.
Во-вторых, предполагается, что такие рецепторы, как SR-A или LRP, представляют собой сильно вытянутые молекулы (~40–50 нм). Кроме того, некоторые из хорошо охарактеризованных опсонинов для апоптотических клеток чрезвычайно велики. Например, C1q составляет примерно 30 нм, а TSP-1 составляет 40–50 нм. Вместе с относительно большими встречными рецепторами для этих опсонинов [e.g., SCARF1 для C1q (107) и α v β 3 для TSP], предсказанное межмембранное контактное расстояние, вероятно, будет несовместимо с таковым для BAI-1, α v β 5 /MFG-E8 , или Mer/Protein S. Одной из возможностей является то, что эти удлиненные структуры эффективны при начальном захвате апоптотических мишеней, облегчая последующее вовлечение пар рецептор/контррецептор, которые охватывают меньшее межмембранное расстояние и находятся под влиянием электростатического отталкивания между клетками.
Вместе с относительно большими встречными рецепторами для этих опсонинов [e.g., SCARF1 для C1q (107) и α v β 3 для TSP], предсказанное межмембранное контактное расстояние, вероятно, будет несовместимо с таковым для BAI-1, α v β 5 /MFG-E8 , или Mer/Protein S. Одной из возможностей является то, что эти удлиненные структуры эффективны при начальном захвате апоптотических мишеней, облегчая последующее вовлечение пар рецептор/контррецептор, которые охватывают меньшее межмембранное расстояние и находятся под влиянием электростатического отталкивания между клетками.
В-третьих, рецепторы семейства Tyro3/Axl/Mer, BAI-1 и интегрин α v β 5 способны инициировать внутриклеточную передачу сигналов, которая контролирует интернализацию частиц. Если есть параллели с образованием иммунологических синапсов, можно ожидать, что эти рецепторы локализуются в центре контактной зоны. Однако межмембранное расстояние ТАМ и интегринов для взаимодействия с их контррецепторами (~30 нм) значительно больше, чем у иммунорецепторов, ответственных за передачу сигналов во время установления иммунологического синапса (~14 нм для класса TcR/MHC). II и костимулирующие рецепторы, такие как CD80/CD28).Мы могли бы предположить, что принципы организации иммунных и эффероцитарных синапсов будут различаться из-за различных требований к распространению сигнала после родственного взаимодействия рецепторов. В иммунном синапсе доставка сигнала, контролирующего пролиферацию Т-клеток или уничтожение клеток-мишеней, требует поддержания межклеточной адгезии и перераспределения антиген-специфических молекул распознавания в зону контакта. Напротив, эффероцитарный синапс, вероятно, является более динамичной структурой, которая облегчает связывание частиц и делает возможной динамическую регуляцию организации цитоскелета по мере интернализации частиц.Как обсуждалось ранее, локализация рецепторов, участвующих в фагоцитозе, может включать начальное связывание, опосредованное более крупными молекулами, как необходимое условие для вовлечения меньших PtdSer-связывающих рецепторов. Механизмы исключения фосфатаз из эффероцитарного синапса также могут различаться.
II и костимулирующие рецепторы, такие как CD80/CD28).Мы могли бы предположить, что принципы организации иммунных и эффероцитарных синапсов будут различаться из-за различных требований к распространению сигнала после родственного взаимодействия рецепторов. В иммунном синапсе доставка сигнала, контролирующего пролиферацию Т-клеток или уничтожение клеток-мишеней, требует поддержания межклеточной адгезии и перераспределения антиген-специфических молекул распознавания в зону контакта. Напротив, эффероцитарный синапс, вероятно, является более динамичной структурой, которая облегчает связывание частиц и делает возможной динамическую регуляцию организации цитоскелета по мере интернализации частиц.Как обсуждалось ранее, локализация рецепторов, участвующих в фагоцитозе, может включать начальное связывание, опосредованное более крупными молекулами, как необходимое условие для вовлечения меньших PtdSer-связывающих рецепторов. Механизмы исключения фосфатаз из эффероцитарного синапса также могут различаться.
Наконец, хотя фагоцитоз интактных мишеней апоптоза легко наблюдается in vitro , плотное прилегание плазматических мембран фагоцитов и мишеней апоптоза в определенных тканях in vivo может потребовать механистически отличного процесса клиренса.Напр., в сетчатке клиренс наружных сегментов фоторецепторов клетками пигментного эпителия сетчатки уподоблен фагоцитарной «обрезке» наружных сегментов фоторецепторов (108). Молекулярная основа удаления наружных сегментов фоторецепторов клетками пигментного эпителия сетчатки включает Mer, Pros1 и Gas6, а также интегрин α v β 5 (108–110). Электронно-микроскопический анализ архитектуры клеток сетчатки показывает чрезвычайно тесный контакт между клетками РПЭ и фоторецепторами (111).Исследования Elegant in vivo показали суточное воздействие PtdSer локализованным образом, что затем запускает «защемление» дистальных концов фоторецепторов соседними клетками RPE (112). Воздействие PtdSer на жизнеспособные фоторецепторы может иметь некоторые параллели с процессом, называемым фагоптозом (113), при котором жизнеспособные клетки распознаются фагоцитами. Однако во время фагоптоза распознавание запускает апоптоз в клетке-мишени (114). Хотя сходные молекулярные пути участвуют в распознавании воздействия PtdSer на жизнеспособные клетки (например,g., MFG-E8, стабилины, интегрины α v и Mer) (115), задействованные события межклеточной коммуникации, вероятно, отличаются от тех, которые необходимы для эффероцитоза.
Однако во время фагоптоза распознавание запускает апоптоз в клетке-мишени (114). Хотя сходные молекулярные пути участвуют в распознавании воздействия PtdSer на жизнеспособные клетки (например,g., MFG-E8, стабилины, интегрины α v и Mer) (115), задействованные события межклеточной коммуникации, вероятно, отличаются от тех, которые необходимы для эффероцитоза.
Таким образом, для эффективного распознавания апоптотических клеток фагоцитами может потребоваться установление эффероцитарного синапса. Кооперативность взаимодействия с рецепторами может способствовать и стабилизировать адгезивные взаимодействия и приводить к сборке сигнальных платформ, которые в конечном счете определяют ответы фагоцитов на связывание и интернализацию апоптотических клеток.
Вклад авторов
ID написал статью и сгенерировал цифры. NB, JM, MV и AR написали статью и критически оценили цифры.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Финансирование
MV признает финансирование от Исследовательского совета по биотехнологии и биологическим наукам (BB/M025160/1).AR частично финансируется Советом медицинских исследований Великобритании (MR/K013386/1). NB признает финансирование от EPSRC и Центра докторантуры MRC в области оптической медицинской визуализации, OPTIMA (EP/L016559/1).
Каталожные номера
4. Savill J, Dransfield I, Gregory C, Haslett C. Взрыв из прошлого: клиренс апоптотических клеток регулирует иммунный ответ. Nat Rev Immunol (2002) 2:965–75. дои: 10.1038/nri957
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
6.Олденборг П.А., Грешам Х.Д., Линдберг Ф.П. CD47-сигнальный регуляторный белок альфа (SIRPalpha) регулирует Fcgamma и фагоцитоз, опосредованный рецептором комплемента. J Exp Med (2001) 193:855–62. doi:10.1084/jem.193.7.855
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
7. Каган В.Е., Гляйсс Б., Тюрина Ю.Ю., Тюрин В.А., Эленстрём-Магнуссон С., Лю С.Х. и соавт. Роль окислительного стресса в апоптозе: окисление и экстернализация фосфатидилсерина необходимы для очистки макрофагами клеток, подвергающихся Fas-опосредованному апоптозу. J Immunol (2002) 169:487–99. doi:10.4049/jиммунол.169.1.487
Каган В.Е., Гляйсс Б., Тюрина Ю.Ю., Тюрин В.А., Эленстрём-Магнуссон С., Лю С.Х. и соавт. Роль окислительного стресса в апоптозе: окисление и экстернализация фосфатидилсерина необходимы для очистки макрофагами клеток, подвергающихся Fas-опосредованному апоптозу. J Immunol (2002) 169:487–99. doi:10.4049/jиммунол.169.1.487
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
8. Chang MK, Bergmark C, Laurila A, Hörkkö S, Han KH, Friedman P, et al. Моноклональные антитела против окисленного липопротеина низкой плотности связываются с апоптозными клетками и ингибируют их фагоцитоз с помощью вызванных макрофагов: доказательство того, что специфичные для окисления эпитопы опосредуют распознавание макрофагов. Proc Natl Acad Sci U S A (1999) 96:6353–8.doi:10.1073/pnas.96.11.6353
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
9. Slater AF, Stefan C, Nobel I, van den Dobbelsteen DJ, Orrenius S. Сигнальные механизмы и окислительный стресс при апоптозе. Toxicol Lett (1995) 8(2–83):149–53. дои: 10.1016/0378-4274(95)03474-9
Toxicol Lett (1995) 8(2–83):149–53. дои: 10.1016/0378-4274(95)03474-9
Полнотекстовая перекрестная ссылка | Академия Google
10. Моррис Р.Г., Харгривз А.Д., Дюваль Э., Уилли А.Х. Гормон-индуцированная гибель клеток. 2. Поверхностные изменения тимоцитов, подвергающихся апоптозу. Am J Pathol (1984) 115:426–36.
Реферат PubMed | Академия Google
12. Martin SJ, Reutelingsperger CP, McGahon AJ, Rader JA, van Schie RC, LaFace DM, et al. Раннее перераспределение фосфатидилсерина плазматической мембраны является общей чертой апоптоза независимо от инициирующего стимула: ингибирование сверхэкспрессией Bcl-2 и Abl. J Exp Med (1995) 182:1545–56. doi:10.1084/jem.182.5.1545
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
13.Фадок В.А., Сэвилл Дж.С., Хаслетт С., Браттон Д.Л., Доэрти Д.Е., Кэмпбелл П.А. и соавт. Различные популяции макрофагов используют либо рецептор витронектина, либо рецептор фосфатидилсерина для распознавания и удаления апоптотических клеток. J Immunol (1992) 149:4029–35.
J Immunol (1992) 149:4029–35.
Реферат PubMed | Академия Google
14. МакЭвой Л., Уильямсон П., Шлегель Р.А. Мембранная фосфолипидная асимметрия как детерминанта узнавания эритроцитов макрофагами. Proc Natl Acad Sci U S A (1986) 83:3311–5.doi:10.1073/pnas.83.10.3311
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
15. Suzuki J, Denning DP, Imanishi E, Horvitz HR, Nagata S. Белок 8, родственный Xk, и CED-8 способствуют воздействию фосфатидилсерина в апоптотических клетках. Наука (2013) 341:403–6. doi:10.1126/наука.1236758
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
16. Yeung T, Gilbert GE, Shi J, Silvius J, Kapus A, Grinstein S. Мембранный фосфатидилсерин регулирует поверхностный заряд и локализацию белка. Наука (2008) 319:210–3. doi:10.1126/наука.1152066
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
17. Park D, Tosello-Trampont AC, Elliott MR, Lu M, Haney LB, Ma Z, et al. BAI1 представляет собой поглощающий рецептор для апоптотических клеток выше модуля ELMO/Dock180/Rac. Природа (2007) 450:430–4. дои: 10.1038/природа06329
BAI1 представляет собой поглощающий рецептор для апоптотических клеток выше модуля ELMO/Dock180/Rac. Природа (2007) 450:430–4. дои: 10.1038/природа06329
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
18. Park SY, Jung MY, Kim HJ, Lee SJ, Kim SY, Lee BH, et al.Быстрое удаление клеточных трупов с помощью стабилина-2, мембранного фосфатидилсеринового рецептора. Cell Death Differ (2008) 15:192–201. дои: 10.1038/sj.cdd.4402242
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
19. Wang X, Li W, Zhao D, Liu B, Shi Y, Chen B, et al. Caenorhabditis elegans Транстиретиноподобный белок TTR-52 опосредует распознавание апоптотических клеток фагоцитарным рецептором CED-1. Nat Cell Biol (2010) 12:655–64. дои: 10.1038/ncb2068
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
20.Андерсен М.Х., Граверсен Х., Федосов С.Н., Петерсен Т.Е., Расмуссен Дж. Т. Функциональный анализ двух клеточных связывающих доменов бычьего лактадгерина. Биохимия (Москва) (2000) 39:6200–6. дои: 10.1021/bi992221r
Т. Функциональный анализ двух клеточных связывающих доменов бычьего лактадгерина. Биохимия (Москва) (2000) 39:6200–6. дои: 10.1021/bi992221r
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
21. Ханаяма Р., Танака М., Мива К., Шинохара А., Ивамацу А., Нагата С. Идентификация фактора, связывающего апоптотические клетки с фагоцитами. Природа (2002) 417:182–7. дои: 10.1038/417182a
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
22.Stitt TN, Conn G, Gore M, Lai C, Bruno J, Radziejewski C, et al. Белок фактора антикоагуляции S и его аналог Gas6 являются лигандами семейства рецепторных тирозинкиназ Tyro 3/Axl. Cell (1995) 80:661–70.
Реферат PubMed | Академия Google
23. Païdassi H, Tacnet-Delorme P, Garlatti V, Darnault C, Ghebrehiwet B, Gaboriaud C, et al. C1q связывает фосфатидилсерин и, вероятно, действует как мультилигандная мостиковая молекула при распознавании апоптотических клеток. J Immunol (2008) 180:2329–38.doi:10.4049/jиммунол.180.4.2329
J Immunol (2008) 180:2329–38.doi:10.4049/jиммунол.180.4.2329
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
24. Беверс Э.М., Комфуриус П., Звал Р.Ф. Изменения в распределении фосфолипидов мембран при активации тромбоцитов. Biochim Biophys Acta (1983) 736:57–66. дои: 10.1016/0005-2736(83)
-4Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
25. Диллон С.Р., Манчини М., Розен А., Шлиссель М.С. Аннексин V связывается с жизнеспособными В-клетками и колокализуется с маркером липидных рафтов при активации В-клеточного рецептора. J Immunol (2000) 164:1322–32. doi:10.4049/jиммунол.164.3.1322
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
26. Fischer K, Voelkl S, Berger J, Andreesen R, Pomorski T, Mackensen A. Распознавание антигена вызывает воздействие фосфатидилсерина на клеточную поверхность CD8+ T-клеток человека. Кровь (2006) 108:4094–101. дои: 10.1182/кровь-2006-03-011742
дои: 10.1182/кровь-2006-03-011742
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
28. Tietjen GT, Gong Z, Chen CH, Vargas E, Crooks JE, Cao KD, et al.Молекулярный механизм дифференциального распознавания мембранного фосфатидилсерина иммунорегуляторным рецептором Tim4. Proc Natl Acad Sci U S A (2014) 111:E1463–72. doi:10.1073/pnas.1320174111
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
29. Frasch SC, Berry KZ, Fernandez-Boyanapalli R, Jin HS, Leslie C, Henson PM, et al. Зависимое от НАДФН-оксидазы образование лизофосфатидилсерина усиливает клиренс активированных и умирающих нейтрофилов через G2A. J Biol Chem (2008) 283:33736–49. дои: 10.1074/jbc.M807047200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
30. Цай Р.К., Дишер Д.Е. Ингибирование «само» поглощения посредством деактивации миозина-II в фагоцитарном синапсе между клетками человека. J Cell Biol (2008) 180:989–1003. дои: 10.1083/jcb.200708043
дои: 10.1083/jcb.200708043
Полнотекстовая перекрестная ссылка | Академия Google
32. Лоуренс Д.В., Кинг С.Б., Фрейзер В.А., Кениг Дж.М. Снижение экспрессии CD47 во время спонтанного апоптоза нацелено на нейтрофилы для фагоцитоза макрофагами, происходящими из моноцитов. Early Hum Dev (2009) 85: 659–63. doi:10.1016/j.earlhumdev.2009.09.005
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
33. Lv Z, Bian Z, Shi L, Niu S, Ha B, Tremblay A, et al. Потеря образования кластеров CD47 на клеточной поверхности и авидности связывания с SIRPα облегчает апоптотический клиренс клеток макрофагами. J Immunol (2015) 195:661–71. doi:10.4049/jиммунол.1401719
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
34.Gardai SJ, Xiao YQ, Dickinson M, Nick JA, Voelker DR, Greene KE, et al. Связывая SIRP-альфа или кальретикулин/CD91, легочные коллектины действуют как наблюдательные молекулы с двойной функцией, подавляя или усиливая воспаление. Cell (2003) 115:13–23. дои: 10.1016/S0092-8674(03)00758-X
Cell (2003) 115:13–23. дои: 10.1016/S0092-8674(03)00758-X
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
35. Savill JS, Henson PM, Haslett C. Фагоцитоз нейтрофилов стареющего человека макрофагами опосредован новым «заряд-чувствительным» механизмом распознавания. J Clin Invest (1989) 84:1518–27. дои: 10.1172/JCI114328
Полнотекстовая перекрестная ссылка | Академия Google
37. Hart SP, Ross JA, Ross K, Haslett C, Dransfield I. Молекулярная характеристика поверхности апоптотических нейтрофилов: последствия функционального подавления и распознавания фагоцитами. Cell Death Differ (2000) 7:493–503. doi:10.1038/sj.cdd.4400680
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
38. Franz S, Frey B, Sheriff A, Gaipl US, Beer A, Voll RE, et al.Лектины обнаруживают изменения статуса гликозилирования компонентов плазматической мембраны во время позднего апоптоза. Цитометрия A (2006) 69:230–9. doi:10.1002/cyto.a.20206
doi:10.1002/cyto.a.20206
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
39. Meesmann HM, Fehr EM, Kierschke S, Herrmann M, Bilyy R, Heyder P, et al. Уменьшение остатков сиаловой кислоты как сигнал еды на поверхности апоптотических лимфоцитов. J Cell Sci (2010) 123:3347–56. дои: 10.1242/jcs.066696
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
40.Джонс Дж., Морган Б.П. Апоптоз связан со сниженной экспрессией регуляторных молекул комплемента, молекул адгезии и других рецепторов на полиморфноядерных лейкоцитах: функциональное значение и роль в воспалении. Immunology (1995) 86:651–60.
Реферат PubMed | Академия Google
41. Takizawa F, Tsuji S, Nagasawa S. Усиление фагоцитоза макрофагов при отложении iC3b на апоптотических клетках. FEBS Lett (1996) 397: 269–72. дои: 10.1016/S0014-5793(96)01197-0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
42. Огден К.А., деКателино А., Хоффманн П.Р., Браттон Д., Гебрехивет Б., Фадок В.А. и соавт. C1q и связывание маннозы с лектином, взаимодействие с кальретикулином клеточной поверхности и CD91 инициирует макропиноцитоз и поглощение апоптотических клеток. J Exp Med (2001) 194:781–95. doi:10.1084/jem.194.6.781
Огден К.А., деКателино А., Хоффманн П.Р., Браттон Д., Гебрехивет Б., Фадок В.А. и соавт. C1q и связывание маннозы с лектином, взаимодействие с кальретикулином клеточной поверхности и CD91 инициирует макропиноцитоз и поглощение апоптотических клеток. J Exp Med (2001) 194:781–95. doi:10.1084/jem.194.6.781
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
43. Арур С., Уче У.Е., Резаул К., Фонг М., Скрэнтон В., Коуэн А.Е. и соавт. Аннексин I представляет собой эндогенный лиганд, который опосредует апоптотическое поглощение клеток. Dev Cell (2003) 4: 587–98. дои: 10.1016/S1534-5807(03)00090-X
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
44. Uehara H, Shacter E. Самоокисление и олигомеризация белка S на поверхности апоптотических клеток необходимы для опосредованного мертирозинкиназой фагоцитоза апоптотических клеток. J Immunol (2008) 180:2522–30. doi:10.4049/jиммунол.180.4.2522
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
45. Харт С.П., Александр К.М., Дрансфилд И. Иммунные комплексы преимущественно связываются с Fc гамма RIIA (CD32) на апоптотических нейтрофилах, что приводит к усиленному фагоцитозу макрофагами и высвобождению провоспалительных цитокинов. J Immunol (2004) 172:1882–7. doi:10.4049/jиммунол.172.3.1882
Харт С.П., Александр К.М., Дрансфилд И. Иммунные комплексы преимущественно связываются с Fc гамма RIIA (CD32) на апоптотических нейтрофилах, что приводит к усиленному фагоцитозу макрофагами и высвобождению провоспалительных цитокинов. J Immunol (2004) 172:1882–7. doi:10.4049/jиммунол.172.3.1882
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
46. Dransfield I, Buckle AM, Savill JS, McDowall A, Haslett C, Hogg N. Апоптоз нейтрофилов связан со снижением экспрессии CD16 (Fc gamma RIII). J Immunol (1994) 153:1254–63.
Реферат PubMed | Академия Google
47. Dransfield I, Stocks SC, Haslett C. Регуляция экспрессии и функции молекул клеточной адгезии, связанная с апоптозом нейтрофилов. Кровь (1995) 85:3264–73.
Реферат PubMed | Академия Google
48. Нисимори Х., Ширацути Т., Урано Т., Кимура Ю., Киёно К., Тацуми К. и др. Новый специфичный для мозга ген-мишень p53, BAI1, содержащий повторы тромбоспондина типа 1, ингибирует экспериментальный ангиогенез. Онкоген (1997) 15:2145–50. doi:10.1038/sj.onc.1201542
Онкоген (1997) 15:2145–50. doi:10.1038/sj.onc.1201542
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
49. Тозелло-Трампон А.С., Кинчен Дж.М., Бругнера Э., Хейни Л.Б., Хенгартнер М.О., Равичандран К.С. Идентификация двух сигнальных субмодулей в пути CrkII/ELMO/Dock180, регулирующих поглощение апоптотических клеток. Cell Death Differ (2007) 14:963–72. doi:10.1038/sj.cdd.4402094
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
52.Мураками Ю., Тиан Л., Восс О.Х., Маргулис Д.Х., Кржевски К., Колиган Дж.Е. CD300b регулирует фагоцитоз апоптотических клеток посредством узнавания фосфатидилсерина. Cell Death Differ (2014) 21:1746–57. doi:10.1038/cdd.2014.86
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
53. Пак С.Ю., Канг К.Б., Тапа Н., Ким С.Ю., Ли С.Дж., Ким И.С. Потребность в адаптерном белке GULP во время опосредованного стабилином-2 поглощения клеточного трупа. J Biol Chem (2008) 283:10593–600.дои: 10.1074/jbc.M70
J Biol Chem (2008) 283:10593–600.дои: 10.1074/jbc.M70
00
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
54. Su HP, Nakada-Tsukui K, Tosello-Trampont AC, Li Y, Bu G, Henson PM, et al. Взаимодействие CED-6/GULP, адапторного белка, участвующего в поглощении апоптотических клеток, с CED-1 и CD91/белком, родственным рецептору липопротеинов низкой плотности (LRP). J Biol Chem (2002) 277:11772–9. дои: 10.1074/jbc.M109336200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
55.Парк Д., Хохрайтер-Хаффорд А., Равичандран К.С. Рецептор фосфатидилсерина TIM-4 не опосредует прямую передачу сигналов. Curr Biol (2009) 19(4):346–51. doi:10.1016/j.cub.2009.01.042
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
58. Savill J, Hogg N, Ren Y, Haslett C. Тромбоспондин взаимодействует с CD36 и рецептором витронектина в распознавании макрофагами нейтрофилов, подвергающихся апоптозу. J Clin Invest (1992) 90:1513–22. дои: 10.1172/ДЖКИ116019
J Clin Invest (1992) 90:1513–22. дои: 10.1172/ДЖКИ116019
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
59. Рабинович М. Диссоциация фаз прикрепления и поглощения макрофагами. Exp Cell Res (1967) 46:19–28. дои: 10.1016/0014-4827(67)
-3Полнотекстовая перекрестная ссылка | Академия Google
60. Гриффин Ф.М., Гриффин Дж.А., Лейдер Дж.Е., Сильверстайн С.К. Изучение механизма фагоцитоза. I. Требования к кольцевому прикреплению лигандов, связанных с частицами, к специфическим рецепторам на плазматической мембране макрофагов. J Exp Med (1975) 142:1263–82. doi:10.1084/jem.142.5.1263
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
61. Dransfield I, Zagórska A, Lew ED, Michael K, Lemke G. Тирозинкиназа рецептора Mer опосредует как связывание, так и фагоцитоз апоптотических клеток. Cell Death Dis (2015) 6:e1646. doi:10.1038/cddis.2015.18
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
62. Гриффин Ф.М., Маллинакс П.Дж. Влияние дифференцировки in vivo и обработки лимфокином in vitro на подвижность рецепторов С3 мононуклеарных фагоцитов человека и мыши. J Immunol (1985) 135:3394–7.
Гриффин Ф.М., Маллинакс П.Дж. Влияние дифференцировки in vivo и обработки лимфокином in vitro на подвижность рецепторов С3 мононуклеарных фагоцитов человека и мыши. J Immunol (1985) 135:3394–7.
Реферат PubMed | Академия Google
63. Сигал Г., Ли В., Арора П.Д., Макки М., Дауни Г., Маккаллох К.А. Участие актиновых филаментов и интегринов в стадии связывания в фагоцитозе коллагена фибробластами человека. J Cell Sci (2001) 114:119–29.
Реферат PubMed | Академия Google
65. Hart SP, Rossi AG, Haslett C, Dransfield I. Характеристика эффектов перекрестного связывания макрофагов CD44, связанных с повышенным фагоцитозом апоптотических PMN. PLoS One (2012) 7:e33142. doi:10.1371/journal.pone.0033142
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
66. Hart SP, Dougherty GJ, Haslett C, Dransfield I. CD44 регулирует фагоцитоз апоптотических нейтрофильных гранулоцитов, но не апоптотических лимфоцитов макрофагами человека. J Immunol (1997) 159:919–25.
J Immunol (1997) 159:919–25.
Реферат PubMed | Академия Google
67. Flannagan RS, Harrison RE, Yip CM, Jaqaman K, Grinstein S. Динамическое «зондирование» макрофагов необходимо для эффективного захвата фагоцитарных мишеней. J Cell Biol (2010) 191:1205–18. дои: 10.1083/jcb.201007056
Полнотекстовая перекрестная ссылка | Академия Google
69. Kayalar C, Ord T, Testa MP, Zhong LT, Bredesen DE. Расщепление актина бета-превращающим ферментом интерлейкина 1 для обратного ингибирования ДНКазы I. Proc Natl Acad Sci U S A (1996) 93:2234–8. doi:10.1073/pnas.93.5.2234
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
70. Конча Н.О., Хед Дж.Ф., Кетцель М.А., Дедман Дж.Р., Ситон Б.А.Аннексин V образует кальций-зависимые тримерные единицы на фосфолипидных везикулах. FEBS Lett (1992) 314:159–62. дои: 10.1016/0014-5793(92)80964-I
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
71. ван Гендерен Х.О., Кенис Х., Хофстра Л., Нарула Дж., Ройтелингспергер СРМ. Внеклеточный аннексин А5: функции связывания фосфатидилсерина и двумерной кристаллизации. Biochim Biophys Acta (2008) 1783:953–63. doi:10.1016/j.bbamcr.2008.01.030
ван Гендерен Х.О., Кенис Х., Хофстра Л., Нарула Дж., Ройтелингспергер СРМ. Внеклеточный аннексин А5: функции связывания фосфатидилсерина и двумерной кристаллизации. Biochim Biophys Acta (2008) 1783:953–63. doi:10.1016/j.bbamcr.2008.01.030
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
72.Гракуи А., Бромли С.К., Сумен С., Дэвис М.М., Шоу А.С., Аллен П.М. и др. Иммунологический синапс: молекулярная машина, контролирующая активацию Т-клеток. Наука (1999) 285:221–7. doi:10.1126/наука.285.5425.221
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
73. Дастин М.Л., Чакраборти А.К., Шоу А.С. Понимание структуры и функции иммунологического синапса. Колд Спринг Харб Perspect Biol (2010) 2:a002311. doi:10.1101/cshperspect.а002311
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
75. Lopes FB, Bálint Š, Valvo S, Felce JH, Hessel EM, Dustin ML, et al. Мембранные нанокластеры FcγRI отделяются от ингибирующего SIRPα при активации макрофагов человека. J Cell Biol (2017) 216:1123–41. дои: 10.1083/jcb.201608094
J Cell Biol (2017) 216:1123–41. дои: 10.1083/jcb.201608094
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
76. Лин Дж., Курилова С., Скотт Б.Л., Босворт Э., Айверсон Б.Е., Бейли Э.М. и соавт. TIRF-визуализация динамики микрокластеров гамма-рецепторов Fc и передачи сигналов на макрофагах во время фрустрированного фагоцитоза. ВМС Иммунол (2016) 17:5. doi: 10.1186/s12865-016-0143-2
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
77. Леупин О., Зару Р., Ларош Т., Мюллер С., Валитутти С. Исключение CD45 из сигнальной области Т-клеточного рецептора в антиген-стимулированных Т-лимфоцитах. Curr Biol (2000) 10:277–80. дои: 10.1016/S0960-9822(00)00362-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
78. Goodridge HS, Reyes CN, Becker CA, Katsumoto TR, Ma J, Wolf AJ, et al.Активация рецептора врожденного иммунитета dectin-1 при образовании «фагоцитарного синапса». Природа (2011) 472:471–5. дои: 10.1038/природа10071
Природа (2011) 472:471–5. дои: 10.1038/природа10071
Полнотекстовая перекрестная ссылка | Академия Google
79. Freeman SA, Goyette J, Furuya W, Woods EC, Bertozzi CR, Bergmeier W, et al. Интегрины образуют расширяющийся диффузионный барьер, который координирует фагоцитоз. Cell (2016) 164:128–40. doi:10.1016/j.cell.2015.11.048
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
81.Чанг В.Т., Фернандес Р.А., Ганцингер К.А., Ли С.Ф., Зибольд С., Макколл Дж. и др. Инициация передачи сигналов Т-клетками сегрегацией CD45 при «близких контактах». Nat Immunol (2016) 17:574–82. дои: 10.1038/ni.3392
Полнотекстовая перекрестная ссылка | Академия Google
82. Flannagan RS, Canton J, Furuya W, Glogauer M, Grinstein S. Фосфатидилсериновый рецептор TIM4 использует интегрины в качестве корецепторов для осуществления фагоцитоза. Mol Biol Cell (2014) 25:1511–22. doi:10.1091/mbc.E13-04-0212
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
83. Kim S, Park SY, Kim SY, Bae DJ, Pyo JH, Hong M и др. Перекрестная связь между рецепторами поглощения stabilin-2 и integrin αvβ5 организует поглощение эритроцитов, подвергшихся воздействию фосфатидилсерина. Mol Cell Biol (2012) 32:2698–708. doi: 10.1128/MCB.06743-11
Kim S, Park SY, Kim SY, Bae DJ, Pyo JH, Hong M и др. Перекрестная связь между рецепторами поглощения stabilin-2 и integrin αvβ5 организует поглощение эритроцитов, подвергшихся воздействию фосфатидилсерина. Mol Cell Biol (2012) 32:2698–708. doi: 10.1128/MCB.06743-11
Полнотекстовая перекрестная ссылка | Академия Google
84. Ву Й, Сингх С, Джорджеску М-М, Бирге РБ. Роль тирозинкиназы Mer в опосредованном интегрином альфаvbeta5 фагоцитозе апоптотических клеток. J Cell Sci (2005) 118:539–53.дои: 10.1242/jcs.01632
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
85. Вернон-Уилсон Э.Ф., Ораде Ф., Браун С.Б. CD31 способствует бета1-интегрин-зависимому поглощению апоптотических Т-лимфоцитов Jurkat, опсонизированных для фагоцитоза фибронектином. J Leukoc Biol (2006) 79:1260–7. doi:10.1189/jlb.1005571
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
86. McCutcheon JC, Hart SP, Canning M, Ross K, Humphries MJ, Dransfield I. Регуляция макрофагального фагоцитоза апоптотических нейтрофилов за счет адгезии к фибронектину. J Leukoc Biol (1998) 64:600–7.
Регуляция макрофагального фагоцитоза апоптотических нейтрофилов за счет адгезии к фибронектину. J Leukoc Biol (1998) 64:600–7.
Реферат PubMed | Академия Google
87. Киркхэм П.А., Спунер Г., Рахман И., Росси А.Г. Макрофагальный фагоцитоз апоптотических нейтрофилов нарушается матриксными белками, модифицированными сигаретным дымом и продуктами перекисного окисления липидов. Biochem Biophys Res Commun (2004) 318:32–7. doi:10.1016/j.bbrc.2004.04.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
88.Барклай А.Н., Ван ден Берг Т.К. Взаимодействие между сигнальным регуляторным белком альфа (SIRPα) и CD47: структура, функция и терапевтическая мишень. Annu Rev Immunol (2014) 32:25–50. doi:10.1146/annurev-иммунол-032713-120142
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
89. Лиллемейер Б.Ф., Мёртельмайер М.А., Форстнер М.Б., Хуппа Дж.Б., Гроувс Дж.Т., Дэвис М.М. TCR и Lat экспрессируются на отдельных белковых островках на мембранах Т-клеток и соединяются во время активации. Nat Immunol (2009) 11:90–6. дои: 10.1038/ni.1832
Nat Immunol (2009) 11:90–6. дои: 10.1038/ni.1832
Полнотекстовая перекрестная ссылка | Академия Google
90. Арас Д., Букар А.А., Боллигер М.Ф., Нгуен Дж., Солтис С.М., Зюдхоф Т.С. и соавт. Новый эволюционно консервативный домен GPCRs клеточной адгезии обеспечивает аутопротеолиз. EMBO J (2012) 31:1364–78. doi:10.1038/emboj.2012.26
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
92. Xiong JP, Stehle T, Diefenbach B, Zhang R, Dunker R, Scott DL, et al.Кристаллическая структура внеклеточного сегмента интегрина альфа Vbeta3. Наука (2001) 294:339–45. doi:10.1126/наука.1064535
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
93. Chattopadhyay K, Lazar-Molnar E, Yan Q, Rubinstein R, Zhan C, Vigdorovich V, et al. Последовательность, структура, функция, иммунитет: структурная геномика костимуляции. Immunol Rev (2009) 229:356–86. doi:10.1111/j.1600-065X.2009.00778. x
x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
94.Дальбек Б., Смит К.А., Мюллер-Эберхард Х.Дж. Визуализация человеческого C4b-связывающего белка и его комплексов с витамин К-зависимым белком S и белком комплемента C4b. Proc Natl Acad Sci U S A (1983) 80:3461–5. doi:10.1073/pnas.80.11.3461
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
95. Хейринг С., Дальбек Б., Мюллер Ю.А. Распознавание лигандов и гомофильные взаимодействия в Tyro3: структурное понимание семейства тирозинкиназ рецепторов Axl/Tyro3. J Biol Chem (2004) 279:6952–8. дои: 10.1074/jbc.M311750200
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
96. Pratt KP, Shen BW, Takeshima K, Davie EW, Fujikawa K, Stoddard BL. Структура домена С2 фактора VIII человека при разрешении 1,5 Å. Природа (1999) 402:439–42. дои: 10.1038/46601
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
97. Руденко Г., Генри Л., Хендерсон К., Ищенко К., Браун М.С., Гольдштейн Дж.Л., и соавт.Структура внеклеточного домена рецептора ЛПНП при рН эндосомы. Наука (2002) 298:2353–8. doi:10.1126/наука.1078124
Руденко Г., Генри Л., Хендерсон К., Ищенко К., Браун М.С., Гольдштейн Дж.Л., и соавт.Структура внеклеточного домена рецептора ЛПНП при рН эндосомы. Наука (2002) 298:2353–8. doi:10.1126/наука.1078124
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
98. Cao E, Zang X, Ramagopal UA, Mukhopadhaya A, Fedorov A, Fedorov E, et al. Кристаллическая структура Т-клеточного иммуноглобулина муцина-3 обнаруживает галектин-9-независимую лиганд-связывающую поверхность. Иммунитет (2007) 26:311–21. doi:10.1016/j.immuni.2007.01.016
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
99.Кук Р.М., Уилкинсон А.Дж., Барон М., Пасторе А., Таппин М.Дж., Кэмпбелл И.Д. и др. Структура раствора эпидермального фактора роста человека. Природа (1987) 327:339–41. дои: 10.1038/327339a0
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
101. Leahy DJ, Aukhil I, Erickson HP. 2.0 Кристаллическая структура четырехдоменного сегмента фибронектина человека, охватывающего RGD-петлю и область синергизма. Cell (1996) 84:155–64. дои: 10.1016/S0092-8674(00)81002-8
Cell (1996) 84:155–64. дои: 10.1016/S0092-8674(00)81002-8
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
102.Лиллис А.П., Ван Дуйн Л.Б., Мерфи-Ульрих Дж.Е., Стрикленд Д.К. Белок 1, родственный рецептору ЛПНП: уникальные тканеспецифические функции, обнаруженные в исследованиях селективного нокаута генов. Physiol Rev (2008) 88:887–918. doi:10.1152/physrev.00033.2007
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
103. Пайк А.С., Бжозовский А.М., Робертс С.М., Олсен О.Х., Перссон Е. Структура человеческого фактора VIIa и его значение для запуска свертывания крови. Proc Natl Acad Sci U S A (1999) 96:8925–30.doi:10.1073/pnas.96.16.8925
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
104. Tan K, Duquette M, Liu JH, Dong Y, Zhang R, Joachimiak A, et al. Кристаллическая структура TSP-1 типа 1 повторяется: новая многослойная складка и ее биологическое значение. J Cell Biol (2002) 159:373–82. дои: 10.1083/jcb.200206062
J Cell Biol (2002) 159:373–82. дои: 10.1083/jcb.200206062
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
107. Ramirez-Ortiz ZG, Pendergraft WF III, Prasad A, Byrne MH, Iram T, Blanchette CJ, et al.Рецептор-мусорщик SCARF1 опосредует клиренс апоптотических клеток и предотвращает аутоиммунитет. Nat Immunol (2013) 14:917–26. дои: 10.1038/ni.2670
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
108. Burstyn-Cohen T, Lew ED, Través PG, Burrola PG, Hash JC, Lemke G. Генетическое исследование взаимодействия ТАМ-рецептор-лиганд при фагоцитозе пигментных эпителиальных клеток сетчатки. Нейрон (2012) 76:1123–32. doi:10.1016/j.neuron.2012.10.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
109.Финнеманн СК. Роль интегрина альфаvбета5 в регуляции фагоцитоза пигментным эпителием сетчатки. Adv Exp Med Biol (2003) 533:337–42. дои: 10.1007/978-1-4615-0067-4_42
Полнотекстовая перекрестная ссылка | Академия Google
110. Finnemann SC, Bonilha VL, Marmorstein AD, Rodriguez-Boulan E. Фагоцитоз наружных сегментов стержней клетками пигментного эпителия сетчатки требует альфа(v)бета5 интегрина для связывания, но не для интернализации. Proc Natl Acad Sci U S A (1997) 94:12932–7.doi:10.1073/pnas.94.24.12932
Finnemann SC, Bonilha VL, Marmorstein AD, Rodriguez-Boulan E. Фагоцитоз наружных сегментов стержней клетками пигментного эпителия сетчатки требует альфа(v)бета5 интегрина для связывания, но не для интернализации. Proc Natl Acad Sci U S A (1997) 94:12932–7.doi:10.1073/pnas.94.24.12932
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
112. Руджеро Л., Коннор М.П., Чен Дж., Ланген Р., Финнеманн С.К. Дневное локальное воздействие фосфатидилсерина на кончики наружных сегментов палочек в сетчатке мышей дикого типа, но не Itgb5-/- или Mfge8-/-. Proc Natl Acad Sci U S A (2012) 109:8145–8. doi:10.1073/pnas.1121101109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
113. Фрикер М., Олива-Мартин М.Дж., Браун Г.К.Первичный фагоцитоз жизнеспособных нейронов микроглией, активированной LPS или Aβ, зависит от фагоцитарной передачи сигналов кальретикулин/LRP. J Нейровоспаление (2012) 9:196. дои: 10. 1186/1742-2094-9-196
1186/1742-2094-9-196
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
114. Хорник Т.С., Вилалта А., Браун Г.К. Активированная микроглия вызывает обратимый апоптоз клеток феохромоцитомы, индуцируя их клеточную гибель путем фагоцитоза. J Cell Sci (2016) 129:65–79. дои: 10.1242/jcs.174631
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
115. Номура К., Вилальта А., Аллендорф Д.Х., Хорник Т.С., Браун Г.К. Активированная микроглия десиалилатирует и фагоцитирует клетки с помощью нейраминидазы, галектина-3 и мертирозинкиназы. J Immunol (2017) 198:4792–801. doi:10.4049/jиммунол.1502532
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
тестовых седанов Hyundai Sonata, Kia Optima, Toyota Camry и Ford Mondeo
Изменения бизнес-седана 2015 года направлены на улучшение визуального стиля и дизайна салона автомобиля.Kia Optima является прямым конкурентом Ford Mondeo и Opel insignia. В Kia рассчитывают, что с обновлением новой модели конкурировать с этими автомобилями на автомобильном рынке будет намного проще. И так, что же нового предложили нам создатели модели «Оптима»? сравнил два поколения автомобилей бок о бок.
В Kia рассчитывают, что с обновлением новой модели конкурировать с этими автомобилями на автомобильном рынке будет намного проще. И так, что же нового предложили нам создатели модели «Оптима»? сравнил два поколения автомобилей бок о бок.
Хотя новая Оптима позиционируется как рейсталинговая модель, она немного подросла в размерах, да и внешний вид немного изменился.Напомним, что третье поколение было выпущено в 2013 году. Для поддержки продаж Kia решила немного освежить популярный седан, чтобы хотя бы несколько лет удерживать продажи на одном уровне.
Что изменилось во внешности новой Оптимы? При сравнении передних частей двух автомобилей сразу бросается в глаза новая решетка радиатора, которая стала намного агрессивнее. Изменился и передний бампер, который стильно сочетается с новыми фарами головного света.Стоит отметить, что впервые в Оптиме появилась функция адаптивного головного света, направляющего лучи фар в сторону руля.
Сбоку новинка получила длинную хромированную окантовку, которая берет начало от передней части автомобиля и проходит по всему кузову к задней части. Форма новой модели стала более обтекаемой, что должно снизить аэродинамическое сопротивление, что скажется на экономичности и немного прибавит автомобилю мощности.
Правда, стоит отметить, что в новой модели за задним пассажирским боковым стеклом по-прежнему находится большой кусок кузовного элемента, который создает большую слепую зону для обзора водителя.
Но без этого элемента кузова общий стиль автомобиля не выглядел бы столь завершенным. Учитывая тот факт, что «Оптима» оснащена камерой заднего вида, проблема большой невидимой области не кажется такой существенной.
Заметны изменения и в задней части.В предыдущей модели спинка была более плоской и выглядела не слишком стильно. Новая модель выглядит намного современнее. Например, более выраженный дизайн задней двери и новые задние фонари полностью меняют восприятие автомобиля.
Интерьер новой Optima также претерпел серьезные изменения. , но в условиях современных требований покупателей слишком простой дизайн интерьера не приемлем. Именно поэтому Киа решила существенно обновить внутреннее оформление салона.
В новой модели стало намного больше разных кнопок, которые стали удобнее. Например, управление системой отопления и кондиционирования стало намного лучше.
Предыдущая модель получила много критики из-за недостатка места в салоне. Именно поэтому инженеры Киа решили увеличить размеры кузова. Правда, точные характеристики пока не обнародованы. Зато можно с уверенностью сказать, что длина автомобиля увеличилась как минимум на 25мм, что конечно добавит места в салоне.
Кроме того, по словам представителя компании, в новой Optima улучшена шумоизоляция, что сделает поездку в автомобиле более комфортной. Сиденья также получили некоторые доработки. Теперь новые сиденья менее подвержены вибрации, что очень важно при дальних поездках между населенными пунктами.
Итак, автомобиль изготовлен из высокопрочных стальных сплавов, а также в некоторых узлах используется углеродное волокно, что позволило инженерам несколько снизить вес автомобиля.
Благодаря редизайну кузова Kia изменила точки крепления подвески, что изменило всю геометрию шасси. Это улучшает управляемость автомобиля, которая оставляла желать лучшего по сравнению с японскими конкурентами.
В России, к сожалению, скорее всего будут представлены только бензиновые двигатели (2,0 и 2,4 литра). Например, в Англии и других странах новая модель также будет оснащена 1,7-литровым дизельным двигателем.
Цены Точные цены и список комплектаций Kiaпока не раскрывает.Однако новая модель не должна быть намного дороже по сравнению с автомобилями нынешнего поколения. Ориентировочная стоимость новой модели будет начинаться от 1,1 миллиона рублей.
Это как минимум на 50 000-100 000 рублей дешевле новой модели Форд Мондео, у которого за эти деньги не будет такой комплектации, как у Оптимы.
Конечно, с выходом новой модели Оптима будет легче конкурировать с такими автомобилями как и , но, тем не менее, Оптима пока не сможет встать на один уровень с этими моделями.Но, как говорят эксперты, Kia уже вплотную подобралась к своим конкурентам и вскоре может возглавить рынок недорогих семейных седанов на мировой арене. Прогресс, как видите, очевиден.
Опытные автолюбители знают, что всю вторую половину прошлого века рынком управляли американцы. Но, ближе к 2000-м, благодаря резкому техническому прогрессу, европейцы, а затем и азиаты стали постепенно догонять своих конкурентов. На данный момент сложно сказать, кто является лидером мирового рынка, но однозначно уже нет такого большого разрыва между компаниями, как это было раньше.В сегодняшней статье мы сравним Ford Mondeo и Kia Optima – очень знаковые и популярные автомобили, имеющие армии поклонников по всему миру.
Ford Mondeo — известный автомобиль среднего класса, разработанный инженерами европейского подразделения Ford. Как отметили представители компании, модель должна была стать глобальной. Даже его название с французского переводится как «мир». Однако в Северной Америке Mondeo не особо прижился, так как даже по местным меркам он казался слишком дорогим.
Как отметили представители компании, модель должна была стать глобальной. Даже его название с французского переводится как «мир». Однако в Северной Америке Mondeo не особо прижился, так как даже по местным меркам он казался слишком дорогим.
Дебют автомобиля состоялся в 1993 году, а уже через 3 года было представлено второе поколение Mondeo. Затем в 2000, 2007 и 2013 годах компания представила следующие три поколения автомобиля. За свою историю модель получила множество наград и премий, в том числе лучший автомобиль 2001 года и лучший автомобиль в своем классе по версии Top Gear.
Киа Оптима, или как ее еще называют в Европе — Киа Маджентис, была выставлена на всеобщее обозрение в 2000 году. Поскольку модель являлась потомком легендарной Сонаты, она сразу же попала в ТОП по продажам.В 2005 году состоялась премьера «Оптимы 2», в которой был улучшен дизайн и модернизирована система безопасности, которая у предшественника считалась очень слабой.
В 2010 году на автосалоне в Нью-Йорке была представлена модель третьего поколения, получившая новый спортивно-футуристический экстерьер.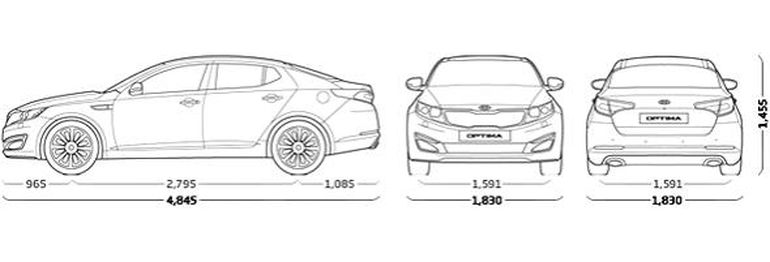 В 2016 году снова в Нью-Йорке дебютировала Optima четвертого поколения, которая, кстати, сейчас находится в Калининграде.
В 2016 году снова в Нью-Йорке дебютировала Optima четвертого поколения, которая, кстати, сейчас находится в Калининграде.
Что лучше — Киа Оптима или Форд Мондео? Учитывая успех американской машины, именно он получает преимущество в этот момент.
Внешний вид
Внешность обоих автомобилей демонстрирует спортивность и прогрессивность, но только с той разницей, что Мондео выглядит солиднее и представительнее, а его соперник агрессивен и целеустремлен.
Впереди у Оптимы можно увидеть широкое лобовое стекло и ровный опускающийся капот. У Mondeo передок оснащен более объемным «передком» и приподнятым выпуклым капотом. На носу «корейца» фирменная решетка радиатора и стильные концептуальные фары головного света.А у Mondeo здесь можно увидеть крупную фальшрадиаторную решетку и высоко расположенные узкие светодиодные фонари. Нижняя часть бампера, особенно воздухозаборники, у обеих моделей очень похожи. А вот противотуманки совсем другие. У Mondeo их роль выполняют крупные, овальные элементы, а у Optima они намного меньше и совершенно незаметны сбоку.
Боковая часть автомобилей имеет много общего. Это и покатая крыша, и аккуратные колесные арки, и плавный профиль.Разве что на боку Mondeo можно заметить тонкую ребристость. Сзади тоже много общего, только бампер у Оптимы выглядит мощнее.
Поскольку во внешности обоих автомобилей много общего, на этом этапе мы присуждаем жеребьевку.
Салон
Заглянув внутрь автомобилей, сразу заметно, что они выполнены в премиум-классе. Однако салоны оформлены в разных стилистических направлениях.Итак, в интерьере Mondeo есть присущая американцам роскошь и хай-тек. В свою очередь салон «Оптима» может предложить достаточно строгий, но в то же время стильный дизайн.
Что касается материалов отделки, то в корейской машине они вроде качественнее и дороже. Однако Mondeo более вместительный и просторный.
Учитывая последний момент и то, что комплектация «американца» богаче, присуждаем Mondeo победу в этом пункте.
Технические характеристики
Учитывая тот факт, что в среднем классе наиболее популярным считается двигатель объемом 2 литра, именно с такими силовыми агрегатами мы выбрали модификации обеих моделей, для сегодняшнего сравнения. Сразу стоит отметить, что обе машины построены на переднеприводной тележке и оснащены шестиступенчатой автоматической коробкой передач.
Теперь вернемся к теме моторов. Оба силовых агрегата оснащены турбинными нагнетателями.Двигатель Mondeo выдает 240 лошадиных сил, а Optima — 245 лошадиных сил. Это, конечно же, сказывается и на показателях эффективности. Время разгона с места до 100 у «корейца» составляет 7,4 с, что на 0,4 с быстрее, чем у соперника. Корейский автомобиль также может похвастаться преимуществом по расходу – против 9 литров у Mondeo.
Что касается габаритов, то ситуация следующая: кузов Mondeo длиннее Optima на 17 мм, но ниже на 7 мм. Колесная база у американского автомобиля намного больше – 2850 мм, против 2805 мм у аналога. Высота клиренса у обеих машин одинакова – 155 мм. Также отметим, что Optima оснащена 18-дюймовыми колесами, а Mondeo — 17-дюймовыми.
Высота клиренса у обеих машин одинакова – 155 мм. Также отметим, что Optima оснащена 18-дюймовыми колесами, а Mondeo — 17-дюймовыми.
Цена
Средняя стоимость Мондео 2017 на отечественном рынке составляет 1 887 000 рублей. обойдется на 57 000 рублей дешевле. Чисто по стоимости лучшим является «Кореец», но, исходя из отзывов покупателей, лучший вариант – Мондео.
Если окна вашей квартиры выходят на дорогу, то откройте окно и прислушайтесь к монотонному гулу ручья.Уберите визг мотоциклов и гул грузовых автомобилей, и у вас останется белый шум, который издают такие машины, как наша. Седаны Hyundai Sonata, Kia Optima, Ford Mondeo и Toyota Camry. Чтобы выбрать один из них, вам даже не нужно ехать к дилеру на пробу. Скучный? Да, но…
Седан Hyundai Sonata имеет долгую и стабильную историю участия в тестах Авторевю. И это история болезни, потому что Сонаты заняли только последние места!
У этой машины затяжной кризис личности.Пару поколений назад, когда еще были Сонаты, все было понятно: большой архаичный седан с удобной подвеской, которая вполне подходила для России. Американский дредноут, поселившийся на чеховских местах. Но затем марка Hyundai взяла курс на Европу — правда, как-то выборочно. Дизайн азиатский, руль пустой, но острый, подвеска трясется, но удара не держит.
Американский дредноут, поселившийся на чеховских местах. Но затем марка Hyundai взяла курс на Европу — правда, как-то выборочно. Дизайн азиатский, руль пустой, но острый, подвеска трясется, но удара не держит.
А вот что такое правильный корейский путь, сестринская марка Kia показывает уже пять лет: с момента своего возрождения в Оптиме в России она неизменно входит в мой шорт-лист рекомендаций к покупке, да и недавняя смена поколений тоже. улучшил свой внешний вид.Мне нравится удобный, просторный салон и центральная консоль с подкладками и прозрачными блоками клавиш, а классические четкие датчики теперь можно найти только в Kia. Наша Оптима в комплектации GT Line — с трехспицевым рулем, толстыми сиденьями, «спортивным» обвесом и ценой 1,7 млн рублей. Красиво, но дорого, ведь техника GT Line ничем не отличается от более простых версий. И это хорошо, ведь если вы никогда раньше не ездили на Оптиме, то вам нужно будет только привыкнуть к габаритам, а в остальном — сесть и поехать. Атмосферный мотор 2.4 (188 л.с.) незаметен на холостых, но дальше тянет бодро и после 4500 об/мин удивляет задорным звуком. Шестиступенчатый «автомат» работает по технологии «стелс», а узнать о его существовании можно, лишь прикоснувшись к подрулевым лепесткам во время движения. Это иногда полезно, потому что спортивная программа силового агрегата в основном меняет отзывчивость педали газа, а не алгоритм работы коробки.
Атмосферный мотор 2.4 (188 л.с.) незаметен на холостых, но дальше тянет бодро и после 4500 об/мин удивляет задорным звуком. Шестиступенчатый «автомат» работает по технологии «стелс», а узнать о его существовании можно, лишь прикоснувшись к подрулевым лепесткам во время движения. Это иногда полезно, потому что спортивная программа силового агрегата в основном меняет отзывчивость педали газа, а не алгоритм работы коробки.
Оптима умеет найти подход и к водителю, и к пассажирам.На руле естественное усилие, отклики в меру резкие, реакции ожидаемые. А если что и случается, то под сбросом газа плавно уходит в занос. При этом плавность хода, особенно с поправкой на 18-дюймовые колеса, достойная: хотя машина и замечает мелкие трещинки и вплавившиеся в асфальт камешки, к короткой волне ведет себя спокойно. И вы перестанете ненавидеть «лежачих полицейских»: Kia успешно берет на себя удары сжатия подвески и не стучит на отбой.А если вам нужно еще больше комфорта, то выбирайте 17-дюймовые диски и тогда вы получаете. .. Hyundai Sonata Sedan!
.. Hyundai Sonata Sedan!
Наконец, Hyundai перестал изобретать велосипед и просто сделал все, как это сделала Kia. Да, есть небольшая разница в настройках шасси: «Соната» чуть тише во всем, усилий на руль в околонулевой зоне немного, реакции более выдержанные. В общем то что нужно для автомобиля такого класса. Еще бы уменьшить внимание к микропрофилю и мелким резким неровностям.Ведь в остальном плавность хода у Сонаты очень хорошая. А на «лежачих полицейских» и вовсе стандарт! Подвеска Hyundai вообще очень хорошо справляется с волнами любой длины и прилично сглаживает неровные стыки — от былого шаткого короткого хода ни следа.
Только слышны отголоски прошлых Сонат: уровень шумоизоляции пока не европейский. Особенно стараются недорогие шины Hankook Kinergy GT, которые гудят на любом асфальте, кроме собянинского, и громко шлепают по щелям.И так же, как и в Киа, двигатель бодрит при разгоне, хотя в окружении Сонаты это менее уместно. Сам силовой агрегат точно такой же, хотя стартовый рывок у Hyundai более быстрый: Sonata достигает 100 км/ч за 8,9 секунды против 9,5 у Kia. Но в целом Оптима и Соната — это пара современных смартфонов среднего класса, которые отличаются только штрихами в электронной оболочке.
Но в целом Оптима и Соната — это пара современных смартфонов среднего класса, которые отличаются только штрихами в электронной оболочке.
Дизайн — тут пока Hyundai остается собой, то есть на любителя.Хотя хромированный пояс, проходящий через весь борт, ностальгически напоминает декор мебельных стенок времен СССР. А лицо Сонаты почему-то сзади: светодиодный уголок габаритных огней воспринимаешь как самую запоминающуюся черту.
Салон чуть пышнее, чем у Киа, центральная консоль вздулась, дисплей оброс дефлекторами, много серебристых кнопок и синих светодиодов — эта Соната оправдывает ожидания любителей Hyundai.Передние сиденья широкие, без претензий на близость пассажира к водителю. А сзади достаточно места для близких отношений с кем угодно, только осторожно перешагните через широкий порог, не прикрытый дверью.
Итак, Соната сейчас не хуже Оптимы, но в большинстве комплектаций все же дороже. Это значит, что в пяти случаях из десяти покупатель этого класса выберет… Camry!
Это настоящий рыночный медоед: он может быть самым старым, самым страшным, самым обременительным в обслуживании и самым дорогим, но все равно останется бестселлером. Недавний рестайлинг сделал лицо менее уродливым, а интерьер вернулся в девяностые. Только в седане Chevrolet Evanda в лучшие годы было столько пластика под дерево. Обновленные датчики досадны — чтобы поднять себе настроение, я специально пересел с новой Камри 2012 года выпуска с ее богатым выбором стрелочных индикаторов (есть даже средний расход топлива!). Зато Camry стала первым серийным автомобилем с установленным мультимедийным приложением Яндекс.Авто. По сути, это привычные для смартфона программы «Карты», «Погода» и «Музыка» от Яндекса, которые сразу интегрированы в автомобиль.Правда, все это доступно только в пакете «Эксклюзив» за 1,7 млн рублей, поэтому выгоднее купить Камри попроще и отдельно приобрести какой-нибудь мультимедийный центр с десятидюймовым экраном на Android: серьезных преимуществ в Работа фирменного модуля.
Недавний рестайлинг сделал лицо менее уродливым, а интерьер вернулся в девяностые. Только в седане Chevrolet Evanda в лучшие годы было столько пластика под дерево. Обновленные датчики досадны — чтобы поднять себе настроение, я специально пересел с новой Камри 2012 года выпуска с ее богатым выбором стрелочных индикаторов (есть даже средний расход топлива!). Зато Camry стала первым серийным автомобилем с установленным мультимедийным приложением Яндекс.Авто. По сути, это привычные для смартфона программы «Карты», «Погода» и «Музыка» от Яндекса, которые сразу интегрированы в автомобиль.Правда, все это доступно только в пакете «Эксклюзив» за 1,7 млн рублей, поэтому выгоднее купить Камри попроще и отдельно приобрести какой-нибудь мультимедийный центр с десятидюймовым экраном на Android: серьезных преимуществ в Работа фирменного модуля.
Эти автомобили имеют качественный комфортный салон и уникальный современный внешний вид. Обе машины лидируют в своем классе среди аналогичных автомобилей.
Оба автомобиля построены по европейским стандартам и предназначены для автомобильного рынка Северной Америки.Какой из этих автомобилей лучше подходит для отечественных клиентов и дорог? Давайте вместе посчитаем.
В американской модели изменена передняя часть, применена новая решетка радиатора и другие детали кузова. Сама решетка имеет форму шестиугольника и похожа на Aston Martin. В целом дизайнеры сделали внешний вид без лишних раздражающих деталей. В целом дизайн экстерьера получился достаточно пламенным и ярким.
Внешний вид корейского автомобиля выглядит спортивно: очертания кузова с обтекаемыми линиями, оригинальные бамперы, «противотуманки» в форме треугольника.В целом внешность Киа Оптима, как и многих марок концерна, выглядит очень молодежно, со спортивными чертами и строгими линиями. Линия подоконника проходит высоко.
Салон Ford Mondeo и Kia Rio
Несмотря на то, что у американца низкая крыша, попасть внутрь машины можно без проблем — места и над головой, и в ногах есть, и спереди, и сзади. В остальном интерьер довольно простой — серые, дешевые пластиковые детали консоли в центре, простые клавиши и много жесткого пластика.Все кресла также простенькие с плохой боковой поддержкой, а спинки набиты очень мягким материалом. Кожаная обивка также проста и недорога.
В остальном интерьер довольно простой — серые, дешевые пластиковые детали консоли в центре, простые клавиши и много жесткого пластика.Все кресла также простенькие с плохой боковой поддержкой, а спинки набиты очень мягким материалом. Кожаная обивка также проста и недорога.
Экран мультимедийной системы расположен неудачно — слишком сильно бликует на ярком солнце. К достоинствам можно отнести задний диван — места хватает во всех положениях, РБ вполне неплох.
Kia Optima в этом плане выглядит лучше. Уникальный дизайн, кожзаменитель на передней панели, обращенной к водителю, качественная мультимедийная система – все выглядит по-европейски.В Kia Optima меньше посадочных мест по сравнению с американским автомобилем. Недостатком задних сидений является низкая крыша и жесткие спинки передних сидений – голова рослых пассажиров упирается в крышу, а колени в спинки передних сидений.
По всем критериям крыша вроде выше по сравнению с американцем, но места для головы не хватает.
Видео
Начало продаж в России
Продажи Киа Оптима в нашей стране начались в начале марта этого года.Продажи обновленного Ford Mondeo в нашей стране начались в начале этого 2016-го года.
Полный комплект
Ford Mondeo:
- Ambient — Двигатель 2,5 л. 149 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 10,3 секунды, максимальная скорость – 204 км/ч, расход: 11,8/6,2/8,2
- В тренде двигатель 2,5 л. 149 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 10,3 секунды, максимальная скорость – 204 км/ч, расход: 11.8/6,2/8,2
- Титаниум — двигатель 2,5 л. 149 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 10,3 секунды, максимальная скорость – 204 км/ч, расход: 11,8/6,2/8,2
- Двигатель 2 л.с. 199 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 8,7 сек, максимальная скорость – 218 км/ч, расход: 11,6/6/8
- Титан + — мотор 2 л.
 с. 199 л. сил, бензин, коробка передач — «автомат», привод передний, разгон — 8.7 секунд, максимальная скорость — 218 км/ч, расход: 11,6/6/8
с. 199 л. сил, бензин, коробка передач — «автомат», привод передний, разгон — 8.7 секунд, максимальная скорость — 218 км/ч, расход: 11,6/6/8 - Двигатель 2 л.с. 240 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 7,9 сек, максимальная скорость – 233 км/ч, расход: 11,6/6/8
 с. 199 л. сил, бензин, коробка передач — «автомат», привод передний, разгон — 8.7 секунд, максимальная скорость — 218 км/ч, расход: 11,6/6/8
с. 199 л. сил, бензин, коробка передач — «автомат», привод передний, разгон — 8.7 секунд, максимальная скорость — 218 км/ч, расход: 11,6/6/8Kia Optima:
- Classic — двигатель мощностью 2 л.с. 150 л. сил, бензин, коробка передач – «механика», привод передний, разгон – 9,6 сек, максимальная скорость – 205 км/ч, расход: 10,4/6,2/7,7
- Комфорт — мотор 2 л.с. 150 л.сил, бензин, коробка передач – «автомат», привод передний, разгон – 10,7 сек, максимальная скорость – 202 км/ч, расход: 11,2/5,9/7,8
- Люкс — мотор 2 литра. 150 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 10,7 сек, максимальная скорость – 202 км/ч, расход: 11,2/5,9/7,8
- Двигатель 2,4 л. 188 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 9,1 сек, максимальная скорость – 210 км/ч, расход: 12,1/6. 2 / 8.3
- Престиж — двигатель 2,4 л. 188 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 9,1 сек, максимальная скорость – 210 км/ч, расход: 12,1/6,2/8,3
- GT-Line — двигатель 2,4 л. 188 л. сил, бензин, коробка передач – «автомат», привод передний, разгон – 9,1 сек, максимальная скорость – 210 км/ч, расход: 12,1/6,2/8,3
- GT — двигатель объемом 2 л. 245 л. сил, бензин, коробка передач — «автомат», привод передний, разгон — 7.4 секунды, максимальная скорость — 240 км/ч, расход: 12,5/6,4/8,5
 2 / 8.3
2 / 8.3Размеры (править)
- Длина форда — 4 м 87,2 сан. Киа — 4 м 85,5 сан.
- Ширина по Форду — 1 м 85,2 сан. Киа — 1 м 86 сан.
- Высота Форда — 1 м 47,8 сан. Киа — 1 м 48,5 сан.
- Клиренс Форд — 12,8 сан. Киа — 15,5 сан.
Цена всех комплектаций
Стоимость Ford Mondeo начинается от 1 млн 350 тыс. руб, заканчивается 2 млн 35 тыс. руб.
Стоимость Киа Оптима начинается от 1 млн 100 тысяч рублей, заканчивается 1 млн 750 тысяч рублей.
Двигатель Ford Mondeo и Kia Optima
Силовые установки Ford Mondeo состоят из 2 двигателей: 2,0 л. и 2,5 литра. Мощность 2 л.с. – 199 и 240 л. сил, мощность 2,5 л. — 149 л. силы. Время разгона от 7,9 до 10,3 с. Средний расход топлива составляет около 6,5 литров по трассе. Максимальная скорость составляет 233 км/ч.
Моторная гамма Киа Оптима состоит из 2-х двигателей — 2 л.с.и 2,4 л. Мощность от 150 до 245 л.с. Время разгона с 7,4 до 9,6 с. Средний расход топлива составляет 6,4 литра. Максимальная скорость составляет 240 км/ч.
Багажник Ford Mondeo и Kia Optima
Багажник американца вмещает 429 литров. Багажник корейца вмещает 510 литров.
Окончательный вывод
Что получилось в результате описанных машин? Комплектация автомобилей неплохая. В корейской модели салон отделан более качественно.А вот в американце — задний ряд просторнее. Что касается стоимости, то Kia Optima несколько дешевле.
Почему автомобилисты выбирают вместительные седаны? Просторные автомобили идеально подходят для поездки на дачу с семьей. Вторая причина заключается в том, что с помощью транспортного средства можно подчеркнуть высокий социальный статус до тех пор, пока не будет собрано достаточно средств для покупки полноценного бизнес-класса. С аргументами разобрались, теперь приступим к выбору конкретной модели.
Вторая причина заключается в том, что с помощью транспортного средства можно подчеркнуть высокий социальный статус до тех пор, пока не будет собрано достаточно средств для покупки полноценного бизнес-класса. С аргументами разобрались, теперь приступим к выбору конкретной модели.
Первыми в списке возможных кандидатов идут Skoda Superb и Volkswagen Passat, но их главный недостаток — высокая цена.На роль большого седана подходит и Toyota Camry, но она очень популярна среди угонщиков, а это очевидный риск: потратить деньги и снова остаться пешеходом через месяц. Выход из этой непростой ситуации есть. «Американский» Ford Mondeo и «корейский» KIA Optima полностью лишены проблем, присущих вышеперечисленным одноклассникам. Какой лучше? Попробуем выяснить.
Внешний вид
Ford Mondeo наделен элегантной «внешностью».Сдержанные линии, классические цвета и строгий дизайн передней части — достойные характеристики для людей, ведущих размеренный образ жизни, предпочитающих больше зарабатывать, чем тратить. Типичный портрет автолюбителя, выбирающего Mondeo: состоятельный семьянин, любящий свою жену и называющий тещу мамой.
Типичный портрет автолюбителя, выбирающего Mondeo: состоятельный семьянин, любящий свою жену и называющий тещу мамой.
KIA Optima не похожа на своего элегантного брата. Автомобиль имеет агрессивную геометрию кузова, суппорта окрашены в ярко-красный цвет. Ее салон — идеальное место для человека, предпочитающего спортивную одежду и яркую обувь.Кто он, водитель Оптимы? Это амбициозный, уверенный в себе мужчина. Этакий мачо.
Хотя представители целевой аудитории, характерные для поклонников каждого из автомобилей, отличаются друг от друга, но владельцы Ford Mondeo все равно завидуют владельцам KIA Optima и наоборот.
Удобство салона
Автомобиль Форд Мондео имеет классический интерьер, даже немного старомодный, хотя в списке материалов отделки присутствует и качественная кожа с замшей.Причина излишней «винтажности» — возраст модели. Американцы выпускают седан в его нынешнем облике уже 5 лет. Но в пользу «Мондео» говорит тот факт, что оснащение салона абсолютно не уступает тому, чем может похвастаться «Оптима». Места для задних пассажиров маловато, а вот багажное отделение гораздо больше, чем у Оптимы.
Места для задних пассажиров маловато, а вот багажное отделение гораздо больше, чем у Оптимы.
С первого взгляда на салон KIA Optima становится понятно, что корейские дизайнеры брали пример с проектов своих немецких коллег.Панель управления мультимедийной системой, не изобилующая лишней информацией, гармонично вписывается в минималистическую центральную консоль. Остальные органы управления расположены так, чтобы водителю было максимально удобно ими пользоваться. Сиденья гораздо больше, чем у «американца», но их обивка явно уступает по качеству.
Ходовые качества
Во время езды Ford Mondeo субъективно превосходит своего корейского одноклассника: отличная управляемость сочетается с хорошей балансировкой подвески.У KIA Optima слишком легкий руль, только на больших скоростях ситуация улучшается. У корейского седана проблемы с быстрым поворотом, поэтому бортовым помощникам приходится включать систему стабилизации. Однако с опытом к особенностям «Оптимы» вырабатывается привычка.
«Мондео» оснащен более мощным двигателем: 199 лошадиных сил против 188 у «Оптимы». Еще одним фактором, говорящим в пользу «американца», является тихий звук двигателя, не раздражающий водителя своим рыком на высоких оборотах.Очевидный плюс Ford Mondeo – плавный ход педали, чем не может похвастаться «кореец». Но есть одно свойство, объединяющее эти два седана, — высокая надежность тормозной системы.
Подведение итогов
Как и следовало ожидать, у обеих машин есть свои плюсы и минусы. Но покупатель в любом случае не пожалеет о своем выборе, чего не скажешь о жене, которая останется без нового смартфона и шубы.
Давайте сравним цены. Ford Mondeo со 199-сильным мотором обойдется в 1 664 000 рублей, а аналогичная версия KIA Optima со 188-сильным мотором на борту обойдется на 75 000 рублей дешевле.«Американец» в максимальной комплектации заставит покупателя раскошелиться на сумму, равную 1 845 000 рублей. «Кореец» в топовой версии GT Line поможет распрощаться с 1 690 000 «деревянных».
Тут следует задуматься: если автолюбитель предпочитает классический стиль, то Ford Mondeo ему подойдет, хотя за «скромность» ему придется выложить почти 150 тысяч и лишить жену престижных подарков. Выбор водителя в дорогом спортивном костюме очевиден — KIA Optima.
Kia Optima 2016 Обзор, технические характеристики, цены и особенности
Салон Kia Optima 2016 модельного года имеет достойное оснащение, начиная с базовой комплектации, что делает его одним из лучших седанов в своем сегменте по соотношению цена-качество.Интерьер получает новый дизайн, а седан приобретает столь необходимую современную привлекательность. Дизайн приборной панели немного ориентирован на водителя со всеми элементами управления в пределах досягаемости водителя. Информационно-развлекательный дисплей также ориентирован на водителя, а новые блоки приборов выглядят лучше, чем раньше. Рулевое колесо получило новый дизайн с тремя спицами с установленными на нем элементами управления для удобства. Передние сиденья предлагают достойный уровень комфорта и просторны даже для высоких взрослых.Задние сиденья тоже просторные, но линия крыши ограничивает пространство над головой для высоких взрослых. В салоне достаточно места для хранения вещей для повседневной практичности, а задние сиденья можно сложить в соотношении 60/40, чтобы увеличить место в багажнике. По умолчанию предлагаемое пространство багажника составляет 15,9 кубических футов.
Передние сиденья предлагают достойный уровень комфорта и просторны даже для высоких взрослых.Задние сиденья тоже просторные, но линия крыши ограничивает пространство над головой для высоких взрослых. В салоне достаточно места для хранения вещей для повседневной практичности, а задние сиденья можно сложить в соотношении 60/40, чтобы увеличить место в багажнике. По умолчанию предлагаемое пространство багажника составляет 15,9 кубических футов.
Модель | Киа Оптима ЛХ | Форд Фьюжн С | База Hyundai Sonata | Ниссан Алтима База |
|---|---|---|---|---|
Вместимость | 5 | 5 | 5 | 5 |
Передний ряд (голова/плечо/нога) (внутри) | 39.8 / 58,1 / 45,5 | 39,2/57,8/44,3 | 40,4/57,9/45,5 | 40,0/56,5/45,0 |
Задний ряд (голова/плечо/нога) (внутри) | 37,8/56,4/35,6 | 37,8/56,9/38,3 | 38. | 37,1/56,4/36,1 |
Грузоподъемность (куб.фут) | 15,9 | 16,0 | 16,3 | 15,4 |
 0 / 56,5 / 35,6
0 / 56,5 / 35,6 Интерьер Ford Fusion выглядит довольно устаревшим по сравнению с салоном Kia Optima и лишен некоторых функций, но предлагает больше места для задних пассажиров по сравнению с Optima.Интерьер Hyundai Sonata имеет некоторое сходство с его родным братом Optima, но ему не хватает ориентированности на водителя. Он предлагает больше места для головы спереди, но его пространство на заднем сиденье также идентично Optima. Nissan Altima имеет стильный дизайн интерьера, но выглядит довольно устаревшим по сравнению с Optima, например Ford Fusion. Но Altima также предлагает более просторный салон, как Fusion. С точки зрения грузового пространства трудно отличить седаны из-за незначительных различий, но статистически Hyundai с 16.3 кубических фута пространства можно назвать победителем.
Некоторые элементы интерьера:
- Аксессуары для полной мощности
- Сиденье водителя с электрорегулировкой по шести параметрам
- Регулируемое по наклону и телескопическому рулевому колесу
- Задние сиденья, складывающиеся в пропорции 60/40
- Настройки памяти драйвера (дополнительно)
- Зеркала с электроприводом складывания
- Рулевое колесо с подогревом
- Кожаная обивка
Kia Optima 2016 года получает 5-дюймовый информационно-развлекательный дисплей с базовой комплектацией, а комплектация EX получает больший 8-дюймовый сенсорный дисплей.Обе системы работают на интерфейсе UVO от Kia, который довольно прост в использовании и имеет удобную компоновку. Плавность интерфейса удивит вас, и он также предлагает почти все основные информационно-развлекательные функции.
Некоторые функции информационно-развлекательной системы:
- 8-дюймовый сенсорный дисплей с информационно-развлекательной системой UVO
- Аудиосистема с 6 динамиками и CD-плеером
- Спутниковое радио
- Bluetooth-телефон и аудиоподключение
- HD-радио
Ручной степлер Rexel Optima 40
ОписаниеСтеплер Rexel Flat Clinch с 1000 скобами в комплекте — RX04813
Степлер Optima 40 Low Force от Rexel — это степлер уникальной конструкции с запатентованным механизмом, который позволяет использовать на 50 % меньше усилий при использовании степлера, чем при использовании традиционных степлеров.
Механизм плоского зажима означает, что при укладывании сшитой бумаги в стопку сшитая сторона не поднимается, так как сзади скобы плоские.
Этот степлер с фронтальной загрузкой Optima 40 имеет привлекательный серый, черный и оранжевый дизайн, который придает ему современный и прочный вид. Он имеет прочную металлическую конструкцию, которая прослужит долгие годы.
Он имеет прочную металлическую конструкцию, которая прослужит долгие годы.
Степлер поставляется со 100 скобами Rexel Optima No.56 (26/6), поэтому вы можете сразу приступить к работе.
Здесь, в Staples, у нас есть огромный выбор скоб самых разных форм и размеров, а также большая коллекция стандартных, тяжелых, красочных и даже электронных степлеров.
Ключевые моменты:
- Цвет: черный, серый и оранжевый
- Тип: Половина полосы
- Размер пакета: Одноместный
- Совместимые скобы: 26/6 и 24/6
- Производители OEM: RX04813
Информация о доставке
БЕСПЛАТНАЯ доставка — когда вы тратите более 33,33 фунтов стерлингов без НДС, без учета марок.
Стандартная поставка — 2,91 фунта стерлингов без НДС 3,49 фунта стерлингов с НДС .
Доставка на следующий день через UPS — 3 фунта стерлингов.90 без НДС
4,68 фунтов стерлингов с НДС (курьер, подписано *).
*Доставка на следующий день не может быть предложена во все места, и иногда поставки могут задерживаться из-за проверки кредитоспособности.
Тяжелая и прямая поставка — 7,49 фунтов стерлингов без НДС 8,99 фунтов стерлингов, включая НДС
Тяжелые и крупногабаритные изделия
Все наши заказы отправляются через курьерские сети и не доставляются бандеролью.Плата за посылку часто увеличивается для заказов весом более 15 кг от всех курьеров, которых мы используем. В результате мы должны отражать это в стоимости доставки, которую мы взимаем с наших клиентов. Заказы, содержащие продукты, вес которых при отправке будет более 20 кг, будут облагаться сбором за доставку больших и больших размеров.
Продукты, которые могут быть затронуты этой посылкой с более высокой скоростью, включают: коробки с бумагой, несколько объемов воды, настольные подставки или мебель, а также объемные упаковки чистящих средств.Плата за перевозку по более высокому тарифу будет указана в вашей корзине как тяжелая или крупногабаритная доставка, стоимость которой составляет 8,95 фунтов стерлингов.